Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
21. Triassic Synapsids in China and India
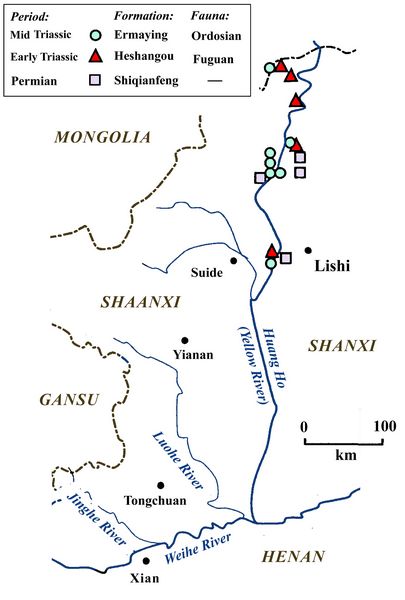
China has three zones of exposed Triassic formations containing vertebrate fossils (fig.1). One of these regions is in the south, located around the boundary of Yunnan and Gouzhou provinces. These are deposits of ancient marine or estuarine formations with an abundance of Triassic marine reptiles, fish, and invertebrates, but few terrestial animals (Benton et al 2013).
 The other two regions are in the north, in the Ordos (Shanxi) and Jungghar
(Xinjiang) Basins. These contain a variety of fossils of synapsids and reptiles,
as well as bony fish, with a much scarcer representation of
labyrinthodont amphibians. In the northeast, the provinces of Shanxi
and Shaanxi straddle the Yellow River, along whose valley many fossil
outcrops are exposed (fig.1). In northwestern China, significant
Triassic exposures occur in the Jungghar basin in Xinjiang province
(Young 1946; Sun 1980). Together, these two regions contain virtually
all of the known land vertebrates or tetrapod fossils from the
Triassic. Lucas (1993) and Sun (1980) summarize the Triassic tetrapod
fauna in northern China, while Benton et al. (2013) review findings
from southern China.
The other two regions are in the north, in the Ordos (Shanxi) and Jungghar
(Xinjiang) Basins. These contain a variety of fossils of synapsids and reptiles,
as well as bony fish, with a much scarcer representation of
labyrinthodont amphibians. In the northeast, the provinces of Shanxi
and Shaanxi straddle the Yellow River, along whose valley many fossil
outcrops are exposed (fig.1). In northwestern China, significant
Triassic exposures occur in the Jungghar basin in Xinjiang province
(Young 1946; Sun 1980). Together, these two regions contain virtually
all of the known land vertebrates or tetrapod fossils from the
Triassic. Lucas (1993) and Sun (1980) summarize the Triassic tetrapod
fauna in northern China, while Benton et al. (2013) review findings
from southern China.Fig.1: Locations of Chinese provinces with Triassic outcrops.
Triassic synapsids and archosaurs in northern China
As late as 1991, no Late Triassic land vertebrate fossils were known in China, although plant fossils definitely existed from that period across northern China (Lucas 1993). This apparent gap in the Triassic vertebrate fossil record was resolved in 1992, when the medium sized archosaur Yonghesuchus sangiensis was found in Upper Triassic strata in Shanxi province (Shao-Chun et al. 2001), as discussed below.
The earliest Mesozoic land vertebrate fauna in China are in western Xinjiang, in the Early Triassic Guodikeng Formation. The faunal assemblage there at the Permian-Triassic boundary is called the Jimsarian fauna (Yang et al. 1992;
 Spencer 1993). This is equivalent to
the Lystrosaurus fauna from South Africa, as discussed by Sun (1980).
Synapsids in the Jimsarian fauna include two dicynodonts,
Lystrosaurus, and the contemporary Dicynodon (called Jimsusaria) ; and
the therocephalian ("beast-headed") carnivore Urumchia (Young
1952).
Therocephalians are advanced synapsids who survived from the Late
Permian into the Early Triassic (ca. 265-245 mya). These were numerous
in South Africa, and spread to Antarctica as well as China, Russia, and
Eastern Europe. Reptiles in the Jimsarian fauna include the procolophon
Santisaurus (Koh 1940); the proterosuchus Chasmatosaurus (Young 1937);
and the protosaur Prolacertoides (Young 1973). All are typical Early
Triassic fauna.
Spencer 1993). This is equivalent to
the Lystrosaurus fauna from South Africa, as discussed by Sun (1980).
Synapsids in the Jimsarian fauna include two dicynodonts,
Lystrosaurus, and the contemporary Dicynodon (called Jimsusaria) ; and
the therocephalian ("beast-headed") carnivore Urumchia (Young
1952).
Therocephalians are advanced synapsids who survived from the Late
Permian into the Early Triassic (ca. 265-245 mya). These were numerous
in South Africa, and spread to Antarctica as well as China, Russia, and
Eastern Europe. Reptiles in the Jimsarian fauna include the procolophon
Santisaurus (Koh 1940); the proterosuchus Chasmatosaurus (Young 1937);
and the protosaur Prolacertoides (Young 1973). All are typical Early
Triassic fauna. The equivalent Early Triassic faunal assemblage from Shanxi and Shaanxi provinces (fig.2) is called the Fugguan fauna, named for the town of Fugu which lies amid the Heshangou Formation (Lucas 1993; Sun 1980). Present are various reptilian taxa, including procolophonids (two species of Eumatabolodon); the protosucherian Xilosuchus, and Fugusuchus, a large Erythrosuchid ("red crocodile"), the largest archosaur predators in the Cygnognathus zone in South Africa. Also from the Fugguan fauna were a few unidentfied remains of labyrinthodont amphibians; and the lungfish Ceratodus ("horned tooth"), similar to examples found in the Bashunchank Formation in Russia.
Fig.2: Late Permian and Triassic vertebrate sites in Shanxi and Shaanxi provinces (after Lucas 1993, fig.3).
Synapsid remains found in Fugguan fauna in Shanxi included the large therocephalian Hazhenia, and smaller therocephalians named Scapulosaurus, also typical of the earliest Triassic of South Africa (Sun 1980; Huttenlocker 2014). Table 2 gives a brief comparison of Early Triassic animal types found in both Xinjiang and Shanxi/Shaanxi.
| Table 1: Comparative Early Triassic fauna in northern China: | ||||
| Region: | Xinjiang | Shanxi/Shaanxi | ||
| Period: | Early Triassic | Early Triassic | ||
| Formation: | Guodikeng | Heshangou | ||
| Faunal assemblage: | Jiimsarian | Fugguan | ||
| Class: | Order: | Group: | Genus: | Genus: |
| Synapsid | Therapsid | Dicynodont |
Lystrosaurus | |
| Synapsid | Therapsid | Dicynodont | Jimsusaria | |
| Synapsid | Therapsid | Therocephalian | Urumchia | Hazhenia |
| Synapsid | Therapsid | Therocephalian | Scapulosaurus | |
| Reptile | Anapsid | Procolophon | Santisaurus | Eumatabolodon |
| Reptile | Diapsid | Protosaurian | Prolacertoides | |
| Reptile | Diapsid | Proterosuchian | Chasmatosaurus | Xilosuchus |
| Reptile | Diapsid | Erythrosuchid | Fugusuchus | |
| Amphibian | Labyrinthodont | Capitosaurid | (unidentified) | (unidentified) |
| Amphibian | Labyrinthodont | Benthosuchidae | (unidentified) | |
| Fish: Ostyichthyes | Sarcoptygerian | Dipnoi (lungish) | Cerotodus | |

Conspicuous by its absence in the Shanxi/Shaanxi fauna is the small synapsid dicynodont Lystrosaurus, otherwise found in Xinjiang, and throughout most of Pangea in the Early Triassic. This discrepancy was reviewed by Sun (1980), who provides a useful summary of the distribution of Lystrosaurus-related fauna in Pangea. Differences in the faunal assemblages between Shanxi and Xinjiang fossil deposits may be linked to geological processes such as variations in micro-plate formation and the physical linkage of different regions of Pangea, something which has been documented for these provinces (Xiong and Coney 1985). These geological factors may have caused differing environments, as well as relative isolation of some areas.
Fig.3: Skull of Parakannemeyeria: A) lateral view, showing position of tusks in maxilla; B) basal view, indicating stapes (after Sun 1960, figs. 2 and 4).
Lower Ermaying Formation
Fossil vertebrates from this component, called the Ordonian fauna by Lucas (1993), consists predominantly of large dicynodonts. Among these is Parakannemeyeria dolichocephala (fig.3), found at the top of the section , together with a variety of Kannemeyeria. The latter animal was widespread in Pangea, found in South Africa, Tanzania, Zambia, Argentina, India, China, and Russia, always in strata at the boundary of the Early and Middle Triassic (Lucas 1993). Kannemeyeria's diagnostic role in this time period (ca. 247-240 mya) is reflected in its use as type species for the Middle Triassic fauna in Xingiangi (table 1). Kannemeyeria is characterized by an long, narrow, and highly projected parietal crest, as well as tusks of larger dimensions in the related Parakannemeyeria (Sun 1960, 1980).
The cynodont Ordosiodon is also represented in the Lower Ermaying Fauna, which is equated to the the Cynognathus Zone of South Africa.
Reptiles from the Lower Ermaying included a moderate sized procolophonid similar to Neoprocolophon, a
 later form which is larger than Procolophon. Several thecodonts
were also recovered from this level, including Fenhosuchus, a primitive
ornithosuchid with armor plates above the neural arches (Sun 1980;
Lucas 1993)
later form which is larger than Procolophon. Several thecodonts
were also recovered from this level, including Fenhosuchus, a primitive
ornithosuchid with armor plates above the neural arches (Sun 1980;
Lucas 1993)Fig.4: Skull of Shansiodon wuhsiangensis, showing large tusks that comprise the two teeth of this dicynodont (after Yuh 1961).
Upper Ermaying Formation
The Ningwuan Fauna from this formation, named after the city of Ningwuan on the Sanggan River in the Ordos Valley, corresponds to the Sinokannemeyeria fauna originally defined by C.C. Young. Young discovered the dicynodont synapsid Sinokannemeyeria pearsoni, the first identification of a Triassic tetrapod in China (Young 1937).
Subsequent work in the Ordos basin took place in the 1950s and 60s under the Chinese Institute of Vertebrate Paleontology and Paleoanthropology (IVPP). This revealed an assemblage of vertebrates that was named the Sinokannemeyeria fauna, thought to correlate with the South African Cynognathus Zone.
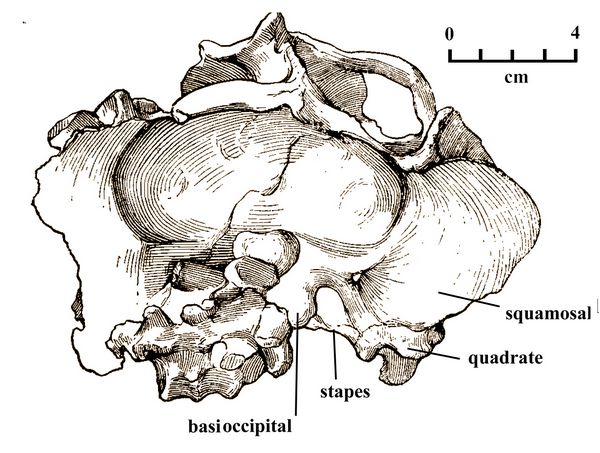
Dicynodonts, representing therapsids who were non-mammalian synapsids, are the most numerous taxa in these late Early Triassic levels, consisting of the three genera Kannemeyeria, Parakannemeyeria, and Shansiodon. The former two are large individuals, while the latter, represented by three different species, is small. One of the Shansiodon species, S. wuhsiangensis, shows the large twin tusks characteristic of the genus (fig.4) . Another, S. wagni (Yuh 1959), showed good preservation of the stapes, a hearing ossicle attached to the quadrate bone in the occipital region at the lower rear of the skull (fig.5).
Fig.5: The skull of Shansiodon wagni viewed from behind, showing the stapes, a small tube-shaped bone used as an ossicle in hearing (after Yuh 1959).
Two Cynodonts (a Therapsid group ancestral to mammals) were also recovered from the Upper Ermaying deposits. One was diagnosed as Sinognathus (fig.6), and another identified as Traversodontoides, a therapsid found most frequently in the Middle Triassic of South America (Young 1959, l974; Sun 1980; Lucas 1993.
Sinognathus belongs to the family Cynognathidae and is regarded as similar to Cynognathus from South
 Africa. Sun (1980), however, notes that, while the South African
cynodont, Cynognathus is a relatively large individual with a skull
that closely resembles a dog, displaying long and narrow post canine
teeth, Sinognathus appears dissimilar. It has an more complex
maxilla or upper jaw area, and more closely resembles Traversodon or
Belesodon.
Africa. Sun (1980), however, notes that, while the South African
cynodont, Cynognathus is a relatively large individual with a skull
that closely resembles a dog, displaying long and narrow post canine
teeth, Sinognathus appears dissimilar. It has an more complex
maxilla or upper jaw area, and more closely resembles Traversodon or
Belesodon.Fig.6: The skull of Sinognathus gracilis, showing its maxilla or upper jaw with a surmaxilla (after Young 1959 fig.3)
Fossils representing the primitive thecodont reptile, Shansisuchus are also abundant, as exemplified by massive fossiliferous block with the vast majority of elements belonging to this genus (Sun 1980). Other thecodont taxa include Fenhosuchus and Wangisuchus. A single skull of the small, temporally diagnostic reptile Procolophonia is also represented in the Upper Ermaying fauna, in the form of Neoprocolophon.
Kelemayi Formation in Xinjiang
The Middle Triassic Kelamayi Formation in Xinjiang contains the Kannemeyeria Fauna (Lucas 1993), which correlates with the entire upper section of the Ermaying Formation exposed along the Yellow River in Shanxi and Shanaxi provinces (Table 2). In more distant regions, according to Sun (1980), the Kelamayi Formation may also be appropriately correlated to equivalent fauna in the Indian Yerrapalli and Dongus System in Russia.
| Table 2: Comparative Early and Middle Triassic fauna in northern China: | |||||
| Region: | Shanxi/Shaanxi | Shanxi/Shaanxi | Xinjiang | ||
| Period: | Middle Triassic | Late Early Triassic | Middle Triassic | ||
| Formation: | Upper Ermaying | Lower Ermaying | Kelemayi | ||
| Faunal assemblage: | Ningwuan | Ordosian | Kannemeyeria | ||
| Class: | Order: | Group: | Genus: | Genus: | Genus: |
| Synapsid | Therapsid | Cynodont | Sinognathus | ||
| Synapsid | Therapsid | Dicynodont |
Parakannemeyeria (3 species) | Parakannemeyeria | Parakannemeyeria |
| Synapsid | Therapsid | Dicynodont | Sinokannemeyeria (2 species) | Kannemayeria | |
| Synapsid | Therapsid | Dicynodont | Shansiodon (3 species) | ||
| Synapsid | Therapsid | Bauriid | Traversodontoides | ||
| Synapsid | Therapsid | Therocephalian | Ordosiodon (3 species) | ||
| Synapsid | Therapsid | Therocephalian | Yikezhaogia | ||
| Reptile | Anapsid | Procolophon | Neoprocolophon | Paotaodon | |
| Reptile | Diapsid | Proterosuchian | Guchengosuchus | ||
| Reptile | Diapsid | Euparkeriid | Wangisuchus | Halazhaisuchus, Turfanosuchus | Turfanosuchus |
| Reptile | Diapsid | Erythysuchian | Shansisuchus (2 species) | Vjushkovia | |
| Reptile | Diapsid | Ornithosuchid | Fenhosuchus | ||
| Amphibian | Labyrinthodont | (unidentified) | (unidentified) | (unidentified) | |
| Fish: Osteichthyes | Semionotid | Sinosemionotus | |||
A Late Triassic Archosaur from Shanxi Province
n 1992 came the first record of Late Triassic tetrapod from China, when fossils of a medium-sized archosaur were found in the southeastern part of the Ordos Basin (Shao-Chun et al. 2001). The fossils included two incomplete skulls with seven neck vertebra and several ribs. After analyzing the fossils, Shao-Chun et al. (2001) identified them as belonging to a new species of Archosaur, which they named Yonghesuchus sangiensis ("crocodilian from Yonghe [county]"). The frequently used "-suchus" designation in Triassic species often indicates crocodile-like dentition in diapsid reptiles who long predate crocodiles, but may be at least distantly ancestral to them. Shao-Chun et al. (2001) describe Yonghesuchus (fig.7) as as the first Archosauriform (Diapsida) known from the Terrestial Upper Triassic of China.

The fossils, embedded in a sandstone block, were found on the north side of Sangbi Creek, about 1.5 km southwest of Sangbi Township, near Lishi in the southeast part of the Ordos Basin. The find was within the Tongchuran Formation, which is higher stratigraphically in the Ordos Basin than the uppermost Ermaying Formation, which dates from the Middle Triassic (Lucas 1993; Sun 1980).
Fig.7: Reconstructed view of the skull of Yonghesuchus sangiensis, with bones labelled (after Shao-Chun et al. 2001, fig.4).
On the basis of stratigraphy, plus associated fossil fish taxa from the Late Triassic found in the same strata as Yonghesuchus, Chao-Chun et al. (2001) identified the fossils as dating from the earliest part of the Late Triassic period. This find thus represents the uppermost stratigraphic record of a land vertebrate in the China Triassic geological formations.
Regarding its phylogeny, Yonghesuchus sangiensis represents a new species of archosaur with some similarities to Turfanosuchus, found in the late Early Triassic, Ermaying formation in Shanxi province, and the Middle Triassic, Kelemayi Formation in Xianjing province (Table 2). Y. sangiensis, however, is considerered more derived than both Turfanosuchus, and the early Late Triassic archosaur Proterochampsa.
Triassic Marine Reptiles in Southern China
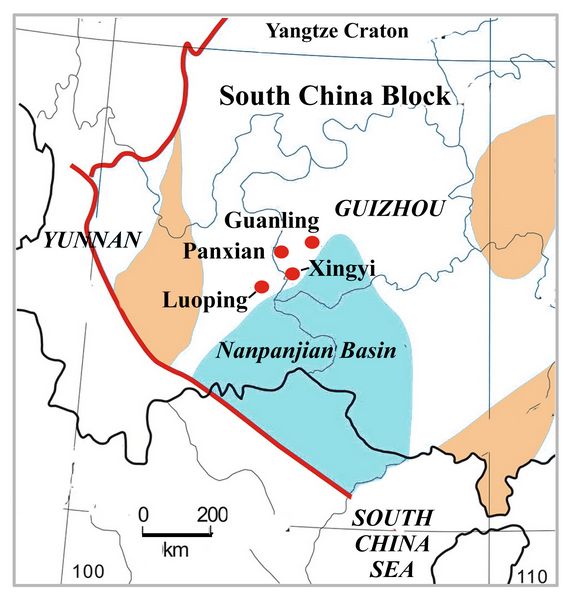
A key zone of fossil-rich geologic formations spanning the Permian-Triassic boundary occurs in the adjacent Yunnan and Ghozou provinces in southern China. This region contains the global standard sections for the Pemian-Triassic boundary, due to both excellect preservation and continuity of fossil assemblages. The South China basin, measuring some 1500 by 3000 km in area, is the zone of rich fossil deposits in continuous sedimentary rock
 strata, serving to record a span of over 70 million years from the Middle Permian to the Late Triassic.
strata, serving to record a span of over 70 million years from the Middle Permian to the Late Triassic. The South China Basin occupies a section of ancient lithosphere known as the South China Block. At the beginning of the Triassic, South China was an island in the Tethys Sea, located just southwest of northern China, at the eastern end of Pangaea (fig.8).
Chinese paleontologists including C.C. Young began intensive work in the Triassic deposits of South China in the 1930s to 70s. Within the past 30 years, several fossil-rich areas have been studied more intensively, with relatively precise chronology now obtained from the strata (Young 1946; Yin et al., 2001; Shen et al., 2011; Benton et al. 2013).
Fig.8: Location of South China in the Tethys sea during the Triassic (after Benton et al. 2013, fig.2A)
Abundant fossil reptiles and fishes have now been excavated from the Triassic of Guizhou Province and neighbouring, eastern parts of Yunnan Province (fig.9). Two of these, the Panxian biota (Hao et al., 2006; Motani et al., 2008; Jiang et al., 2009), and the Luoping biota, date from the Anisian stage of the early Middle Triassic (252-247 mya). A third is the Guanling biota, dating from the from the Carnian stage of the Late Triassic, dated 235-228 mya (Wang et al., 2003; Jiang et al., 2005),
Probably the most spectacular finds have come from the the Luoping biota of the early Middle Triassic. This represents an extensive array of both marine invertebrates and vertebrates. These shallow marine deposits show arthropods as their most abundant fossils, followed by fishes, marine reptiles, bivalves, gastropods, echinoderms, brachiopods, conodonts, foraminifers, and plants (Zhang et al., 2008; Hu et al., 2011).
Nearly 20,000 individual macrofossils have been identified from the Luoping biota in its primary deposits (Benton et al 2013). Recent invertebrate finds include well-preserved belemnoids (cephalopods with bony parts,resembling small squids), hydromedusae, ammonoids, and lingulid brachiopods. The dominant arthropods include crustaceans such as decapods (lobsters and shrimps), isopods, cycloids, and ostracods, tiny shrimp-like animals whose abundance and variation make them used as chronological markers (second only to conodonts). Other Triassic arthropods include millipedes and horseshoe crabs.
Fig.9: Triassic fossil deposits in south China, showing Luoping, Panxian, and Guanling biotas (Benton et al. 2013).
Reptiles from the Luoping biota include medium-sized (1-5 m -long) sea reptiles such as Mixosaurus, Dinocephalosaurus, and Sinosaurosphargis, plus unidentfied archosaurs.
Early Triassic Land Vertebrates in India
India contains regions with a continuous succession of Early, Middle, and Late Triassic sediments, containing well preserved terrestial faunas. In Eastern India, Early Triassic land vertebrates have been identified from the Panchet Formation of the Raniganj coalfield of Damodar Valley (Bandyopadhyay 1999).
Temnospondyl amphibians in the Panchet Formation include the trematosaurid Gonioglyptus, a widespread taxa found from Antarctica to Russia. Very similar trematosaurids are in the Prionolobus beds of the Salt Range, of Dienarian age. Four other temnospondyls are known. These include Indobrachiops, of the Indobrachyopid group, which is restricted to India and Australia. Also found are Lydekkerina, the type genus of the widespread Lyderkkerinid group, found from Antarctica to Russia; Pachygonia of the Captitosaurids, another widespread group found from Antarctica to Russia; and Indobenthosuchus, which appears related to Lydekkerina.
Synapsids from the Panchet Formation include the widespread dicynodont Lystrosaurus, representing tetrapods of lowland habitats living in fluvial and lacustrine settings (Bandyopadhyay 1999). Also found is the cynodont Thrinaxodon, associated with the Lystrosaurus Zone in South Africa, representing the initial phase of the Early Triassic. Table 3 lists Early and Middle Triassic vertebrate fauna from three regions of India.
A herd of Indian Triassic herbivores
Wadiasaurus indicus, a small Middle Triassic dicynodont herbivore about 50 cm long, has been identifed as a kannemeyeriid, so far the only example of this family of therapsid grazers known in India. The Wadiasaurus type specimen is a skull found in an Anisian (
 247-242 mya)
fluvial claystone deposit in the Yerrapalli Formation of the Pranhita-
Godavari valley, about 2 km east-south-east of the town of Yerrapalli.
The genus is named for the Indian geologist Darashaw Nosherwan Wadia
(1883-1969).
247-242 mya)
fluvial claystone deposit in the Yerrapalli Formation of the Pranhita-
Godavari valley, about 2 km east-south-east of the town of Yerrapalli.
The genus is named for the Indian geologist Darashaw Nosherwan Wadia
(1883-1969).More recent findings of a larger sample of Wadiasaurus fossils, in a locality near that of the original type skull, have provided more information on the lower jaw and postcranial skeleton, indicating W. indicus was closely related to Kannemeyeria erithea (Bandyopadhyay 1988). In Wadiasaurus, the parietal crest at the top of the skull is quite specialized, forming two long rounded lobe-like bars separated by a bay. Sexual dimorphism of the species is indicated by maxillary flanges, which in males are triangular, thick and swollen, with stout cylindrical tusks, while similar maxillary flanges of females lack tusks.
Fig.10: Skull of Rechnisaurus.
Interesting behavioral traits have been revealed by a taphonomic study of the bone assemblage found in the fluvial claystones near Yarapalli. These findings show that a herd of Wadiasaurus, including some juveniles and young animals, was trapped in the soft muds of a floodplain and buried in a small area. The herd wa composed only of females, with some juvenile members. Taphonomic and osteological studies tend to indicate that the female individuals of Wadiasaurus lived in herds, whereas the solitary males joined the herds only during the mating seasons (Bandyopadhyay 1999).
A second dicynodont from the Pranhita-Gondavari valley is Reichnisaurus (fig.10). This shows similarities to a South American taxa named Dinodontosaurus, from the Stahlekriid family (Colbert 1984; Bandyopadhyay 1988; Jain 1996).
References:
Anderson and Cruikshank 1978
Bandyopadhyay, S. 1988. A Kannemeyeriid Dicynodont from the Middle Triassic Yerrapalli Formation. Phil. Trans. R. Soc 320 (1198), pp. 185–233
Bandyopadhyay, S. 1999. Gondwana Vertebrate Faunas of India. PINSA 65(8), pp.285-313.
Barberena et al. 1985
Barrett 2000
Benton, M.J. 2005 . When life nearly died: the greatest mass extinction of all time. London: Thames & Hudson
Benton, M.J. et. al. 2013
Chowdhury. T.R. 1970. Two new dicynodonts from the Triassic Yerrapalli Formation of central India. Palaeontology 13, pp.133-144
Colbert, E.H. 1973 The Labyrinthodont-Chilotherium Complex.
Colbert, E.H. and J.W. Kitching 1975 The Triassic reptile Procolophon in Antarctica. American Museum Novitates no. 2566.
Colbert, E.H. 1984
Ezcurra, M.D. 2014 The Osteology of the Basal Archosauromorph Tasmaniosaurus triassicus from the Lower Triassic of Tasmania, Australia . PloS One 9(1)
Hong Youchong, 1985: New discovery of triopsids Notostraca, Early Triassic, Shanxi Province. Journal of Changchun College of Geology = Changchun Dizhi Xueyuan Xuebao 1985(3(41)): 9-12
Jain, S.L. 1996. Aspects of Vertebrate Fossils from Prahita-Godavari Valley with Emphasis on Dinosaur Discoveries. Journal of the Palaentological Society of India 41, pp. 1-16.
Lucas, S.G. 1993 Vertebrate biochronology of the Triassic of China. In Lucas, S.G. and Morales, M. (eds.). The Nonmarine Triassic. New Mexico Museum of Natural History & Science Bulletin. pp. 301–306.
Mazin, J. M. and King, G. M. (1991). The first dicynodont from the Late Permian of Malagasy. Palaeontology 34, pp.837–842.
Shao-Chun, Wu, Liu Jun, and L.I. Jin-Ling 2001. Anatomy of the first Archosauriform (Diapsida) from the Terrestial Upper Triassic of China. Vertebrata Pal Asiatica, vol 10, ppl 251-265
Sun, Ailing 1980 Late Permian and Triassic Terrestial Tetrapods of North China. Vertebrata PalAsiatica Vol XVIII, No.2 April, 1980 pp. 100-110
Surkov, M.V.; and Benton, M.J. 2008 Head kinematics and feeding adaptations of the Permian and Triassic dicynodonts. Journal of Vertebrate Paleontology 28 (4): 1120–1129.
Univ. of Calif. Paleontology website, 2014a Protocolophon.
Wang, R., Xu, S., , Wu, X., Li, C., and S. Wang. 2013. A New Specimen of Shansisuchus shansisuchus Young, 1964 (Diapsida: Archosauriformes) from the Triassic of Shanxi, China. Acta Geologica Sinica (English Edition).
Young, C.C. 1937. On the Triassic Dicynodonts from Shansi. Bull. Geol. Soc. China, Vol.17, Nos. 3-4, pp. 393-412.
Young, C.C., 1957. Neoprocolophon asiaticus, a new Cotylosaurian Reptile first discovered from China. Vertebrata PalAsiatica. Vol. 1, No. 1, pp. 1-8.
Young, C.C., 1959. Note on the first Cynodont from the Sinokannemeyeria fauna in Shansi, China. Vertebrata PalAsiatica. Vol. 1, No. 1, pp. 124-131.
Young, C.C.; 1961. A New Cynodont from Northern Shanxi. Vertebrata PalAsiatica. February 109-113
Young, C.C.; 1964. Pseudosuchians of China, Journal of Paleontology No. 151 #19
Young, C.C.; 1974. A New Traversodon from Jiyuan, Henan. Vertebrata PalAsiatica 12(3): 203-206
Young, C.C.; 1979. A New Late Permian Fauna from Henan", Vertebrata PalAsiatica 17(2), 19-113
Young, C.C., and Yeh, Yangkui; 1963. The First Discovery of the Pareiasauria in China, Vertebrata PalAsiatica 7 (3): 195-214
Xiong, Ji and P.J. Coney l985. Accreted Terranes of China, in Howell, D.G.(ed.), Tectonostratigraphic Terranes of the Circum Pacific Region, pp.349 361,
Glossary