Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
14. Amphibians 2: Reptiliomorphs
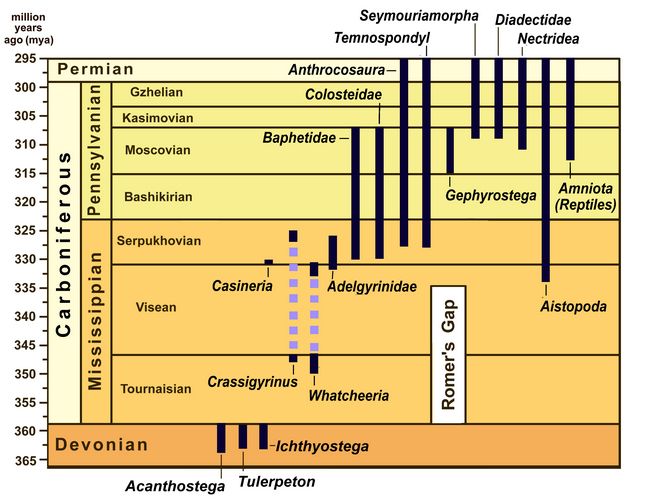
During the Carboniferous era (Mississippian and Pennsylvanian periods), lasting from 358.9 to 298 mya, a centrally important order of Amphibians arose now known as Reptiliomorphs (originally called Anthracosaurs ["coal lizards"], because of the high incidence of their fossils in coal measures; see also below). From the Reptiliomorphs evolved both the Sauropsid reptiles, ancestors of all later reptiles, and the synapsids, who led eventually to early mammals.
Fig.1: Chronological Chart of the Carboniferous era, showing major groupings of amphibians, and placement of Romer's Gap at lower right.
Reptiliomorphs or Anthracosaurs.
Reptiles developed from amphibians in the later part of the Carboniferous era (Late Mississippian and Pennsylvanian periods, about 330-298 mya). The first transitional forms are alternatively called Reptiliomorphs (Säve-Söderbergh 1934; Panchen 1970) or Anthracosaurs (Huxley 1867; Romer 1956; Carroll 1988). These two (largely synonomous) major groupings of amphibians (reptiliomorpha and anthracosauria) are used inconsistently in the literature partly because, when originally defined, they included groups now considered less closely related. There is currently no real consensus on which term is preferable. The name Reptiliomorpha was first used by Gunnar Säve-Söderbergh in 1934 to designate various types of late Paleozoic reptile-like amphibians with labyrinthodont teeth. In 1956 Friedrich von Huene assigned both amphibians and anapsid reptiles to the Reptiliomorpha. This included eight orders, now considered to belong to distinct lineages, whose similarities are the result of convergences or shared primitive features (Palaios 2014d)
Romer (1956), in his widely-read textbooks on vertebrate paleontology, adhered to the term Anthracosauria that T.H. Huxley had originally used in the 1860s, based on fossil findings in coal deposits. In 1970, meanwhile, the German paleontologist Panchen (1970) reverted to Säve-Söderbergh's use of the term Reptiliomorpha. The term Anthracosauria, however, is still in use, both by evolutionary taxonomists who follow Romer (e.g. Carroll 1988 and 2002, and Hildebrand & Goslow 2001), and some cladistic taxonomists (Gauthier et al 1988). In an attempt at clarification, Michael Benton (2000, 2005) made Reptiliomorpha the sister-clade to Batrachomorpha, ancestors of all current amphibians. In this solution, Reptiliomorpha is made into a superorder, and Anthracosauria into an order (Palaios 2014d).
Reptiliomorphs are grouped into various suborders including Embolomeri, Diadectomorpha, Gephyrostega, and Seymouriamorpha, most named for diagnostic or index taxa for each suborder.
Emblomeri.
The Emblomeri, with transitional features of both amphibians and early reptiles, arose in the Late Mississippian period and lasted through the Pennsylvanian.
Protogyrinus
Proterogyrinus ("early tadpole") was a large anthracosaur amphibian from the Serpukhovian stage of the Late Mississippian period (330-323 mya). The type species, P. scheelei, was named by Alfred Romer in 1970, and the taxon was further described by Holmes in 1984. Two species are known, P. scheelei and P. pancheni. Fossils have been found in Scotland, and West Virginia.
Proterogyrinus grew up to 2 1/2 m in length, similar in size and shape to other large amphibians such as Crassigyrinus and Eryops, and showed a mixture of pr
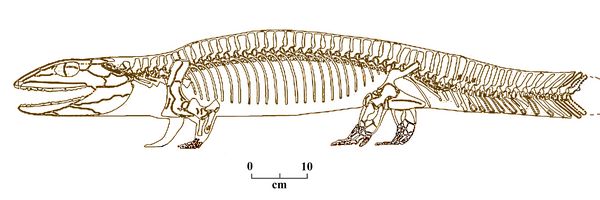 imitive (tetrapod-like) and derived traits.
imitive (tetrapod-like) and derived traits. Proterogyrinus had a semi-elongated skull, taller than that of other early tetrapods such as colosteids and temnospondyls, and resembling those of other embolomeres such as Archeria. Its rounded skull was amphibian, still retaining a fish-like hinged skull, with a hinge at the neck.
Fig.2: Proterogyrinus scheelei (after Romer 1970)
In its postcranial anatomy, Proterogyrinus retained amphibian ankle bones, and had small shoulder girdle bones. Yet it also had reptilian vertebrae, pelvis, humerus, and digits, with both five toes and five fingers. Strong limbs with several fully-ossified ankle and wrist bones allowed Proterogyrinus to walk and hunt on land. Like other reptiliomorphs, such as Seymouria, Proterogyrinus could move further away from water than most amphibians.
However, the presence of lateral line grooves and otic notches which likely held spiracles indicate they retained aquatic, early tetrapod traits. Their long, high tail was used in swimming, and their eye position, high on the skull, suggests a viewpoint just above the water surface.
Diadectomorpha.
More developed reptiliomorph amphibians with additional reptilian features arose in the late Pennsylvanian through Early Permian periods. A suborder of these more advanced forms, called the Diadectomorphs, include the type species Diadectes, Limnoscelis, and Tseajaia.
Diadectes
 Diadectes sideropelicus ("cross-biter of iron clay"), was a
reptiliomorph dating from the Early Permian Cisuralian stage (290–272
mya). It was first found in the Permian red beds of north Texas, in the
Wichita and Clear Fork group near Wichita Falls. The initial
identification was made in 1878 by Copes, who collected extensively
there.
Diadectes sideropelicus ("cross-biter of iron clay"), was a
reptiliomorph dating from the Early Permian Cisuralian stage (290–272
mya). It was first found in the Permian red beds of north Texas, in the
Wichita and Clear Fork group near Wichita Falls. The initial
identification was made in 1878 by Copes, who collected extensively
there.Fig.3: Skull of Diadectes sideropelicus (Yale Peabody Museum; photo: Athena Review.)
The anatomy of Diadectes shows a mixture of reptilian and amphibians traits, combining a robust, reptile-like skeleton with a more primitive, thick-boned skull resembling that of a Seymourian amphibian (fig.3). Its heavy vertebrae and ribs, massive limb girdles and short, robust limbs indicate that D. sideropelicus was a fully terrestrial animal. Diadectes was one of the very earliest herbivorous tetrapods. Its teeth show advanced specializations for herbivorous diet. Eight front teeth are spatulate and peg-like,serving as incisors to clip off mouthfuls of vegetation. The broad, blunt cheek teeth, used for grinding, show extensive wear associated with occlusion.
Limnoscelis
Limnoscelis (fig.4) was a relatively large reptiliomorph carnivore in the Diadectomorph suborder, about two meters in length. The type species Limnoscelis paludis ("marsh-footed swamp dweller") was found by Samuel Williston (1911a) in El Cobre Canyon in the Cutler Formation of New Mexico, dating from the Late Pennsylvanian and Early Permian periods (310-280 mya),
Limnoscelis shows a mixture of reptilian and amphibians traits, combining a robust, reptile-like skeleton with a more primitive, thick-boned skull resembling that of a Seymourian amphibian. The skull of Limnoscelis had a deep premaxilla with robust premaxillary fangs. The maxilla or upper jaw was deeper than in Seymouria, with short teeth which indicate a carnivorous diet. The eye orbit was relatively smaller than
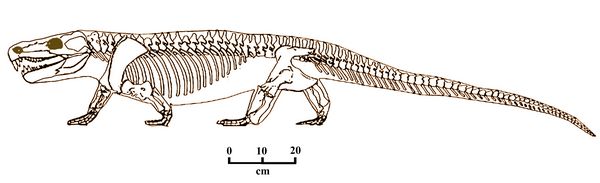 in the related taxa Oedaleops, but in
most other respects, the skulls were similar. Its pectoral and pelvic bones were large and robust, showing a
well-developed mode of walking. Heavy vertebrae and ribs, massive
limb girdles and short, robust limbs indicate that Diadectamorphs were
fully terrestrial animals.
in the related taxa Oedaleops, but in
most other respects, the skulls were similar. Its pectoral and pelvic bones were large and robust, showing a
well-developed mode of walking. Heavy vertebrae and ribs, massive
limb girdles and short, robust limbs indicate that Diadectamorphs were
fully terrestrial animals.Fig.4: Limnoscelis (after Benton 2005).
Other reptilian traits include a larger braincase structure, reptilian jaw muscles, and expanded neural arches.
Its arms and hands showed the primitive tetrapod traits of humerus, radius, ulna, and five digits. The digits lacked claws. The ankle bones, meanwhile, were fused as in other reptile-like amphibians, which would not allow them to use their feet actively in traction.
Gephyrostega
Gephyrostegus bohemicus was a Late Carboniferous, lizard-like reptiliomorph from the Pennsylvanian period (about 310 mya), found at Nyran in the Czech Republic (Jaeckel 1902; Brough and Brough 1967). It is thought to have been derived from a form similar to Silvanerpeton. Gephyrostregus is considered directly ancestral to early reptiles, including Casineria and Cephalerpeton.
Gephyrostegus, with an overall length of about 22 cm., was smaller than other basal tetrapods and slightly smaller than its more primitive sister, Silvanerpeton. The top and sides of the body of Gephyrotegus was covered by tiny circular scales, while on its underside, there were large V-shaped scales. Juveniles had a relatively larger head and orbit (eye socket). In the rear of its skull
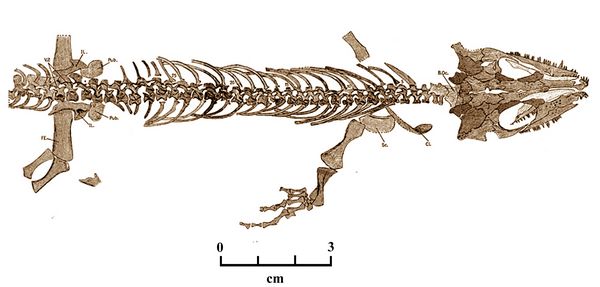 , the otic notch was
reduced. Some postcranial bones of Gephyrostegus were relatively shorter
than those of Silvanerpeton, including the pelvis, and the four
and fifth hand digits. In its ankles, some bones had fused together to
create a larger bone, the astragalus, otherwise found only on reptiles.
Further, an elbow or olecranon process had developed on the ulna, one
of the two lower arm bones. The humerus (upper arm bone) was more also
more slender or gracile.
, the otic notch was
reduced. Some postcranial bones of Gephyrostegus were relatively shorter
than those of Silvanerpeton, including the pelvis, and the four
and fifth hand digits. In its ankles, some bones had fused together to
create a larger bone, the astragalus, otherwise found only on reptiles.
Further, an elbow or olecranon process had developed on the ulna, one
of the two lower arm bones. The humerus (upper arm bone) was more also
more slender or gracile.Fig.5: Gephyrostegus bohemicus (after Brough and Brough 1967, fig.10).
While most early tetrapods lived their lives in water, Gephyrostegus preferred land. Its eggs would have been laid in or near water to prevent dessication because, presumeably, no amniotic membrane or shell surrounded the embryo.
Seymouriamorpha
Seymouriamorpha were a small but widespread sub-order of reptiliomorphs. Many seymouriamorphs were terrestrial or semi-aquatic. However, aquatic larvae bearing external gills and grooves from the lateral line system have been found, making them unquestionably amphibians. The adults were terrestrial. They ranged from lizard-sized creatures (30 cm) to crocodile-sized, 1.5 meter-long, reptile-like animals. The last seymouriamorph became extinct by the end of the Permian period (252 mya).

Seymouriamorphs form into three main groups, Kotlassiidae, Discosauriscidae, and Seymouriidae, a group that includes the best known genus, Seymouria (fig.5), dating from the Early Permian of North America and Europe (298-270 mya).
Seymouria is known from two species, S. baylorensis and S. sanjuanensis. The type species, S. baylorensis, found in Texas, is more robust, while the smaller species, S. sanjuanensis, is more widespread, found in both New Mexico and Germany.
Fig.6: Seymouria baylorensis skeleton (US National Museum of Natural History).
Seymouria were well adapted to life on land, with many reptilian features, resulting in its original identification as a primitive reptile (Williston 1911b; Watson 1919; Romer 1928). Seymouria's body shape resembled that of early reptiles such as captorhinids, with strongly-built limbs and backbone indicating that Seymouria was primarily terrestrial.
In the 1950s, however, fossilized tadpoles were discovered in Discosauriscus, a close relative of Seymouria in the group Seymouriamorpha. This shows that seymouriamorphs had a larval stage which lived in the water, therefore making Seymouria an amphibian closely related to reptiles (Bratislava 1995). Seymouria is an important transitional fossil taxon, documenting the acquisition of reptile-like skeletal features prior to the evolution of the amniotic egg.
To be covered in more detail in the next section, a major difference between these ancient, large amphibians and the first reptiles, who were small in stature, is the amniotic egg which developed in reptiles. Here the embryo is contained in an egg with nutrient fluid and a protective sheath, as opposed to amphibians and fish whose larvae are hatched in open water.
Additional differences developed by reptiles from their amphibian ancestors include stronger legs and girdles, different vertebrae, and stronger jaw muscles (cf. Carroll 1988; Gauthier et al. in Benton, 1988). The reptilian skull was also much stronger. The ancestral amphibians had a relatively weak skull inherited from the fish-like tetrapods.
Later Reptiliomorphs (Permian and Triassic Periods)
Chroniosuchia.
Chroniosuchians, first defined by Kuhn (1970) were a suborder of reptiliomorphs that lived from the Middle Permian to Late Triassic in what is now Eastern Europe, Kyrgyzstan, China and Germany. They were all short limbed, with a strong tail and elongated snout somewhat resembling that of modern crocodiles. The group is traditionally considered to be a suborder of labyrinthodonts. Chroniosuchians likely existed as riverine predators.
Most forms bore a heavy armour of scutes along the back, possibly for protection against land predators. This is illustrated by a species, Chroniosaurus dongusensis from the upper Tatarian to Early Wuchiapingian levels from the Late Permian period in the Orenburg Region of the South Cis-Urals.
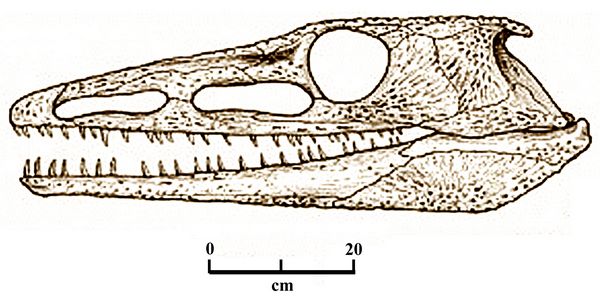 C. dongusensis had diagostic
rows of interlocking bony plates called osteoderms along their backs,
reaching from head to tail (Tverdokhlebova, 1972). These osteoderms
constitute the most frequently found fossilized remain of
Chroniosaurus. The flat osteoderm plates are connected to the neural
arches of vertebra by an extension of bone on their undersurfaces. Each
osteoderm thus is paired with a single vertebra. The front margin
of each osteoderm also has a pair of "anterior wings" that slip into a
notch in the posterior margin of the osteoderm in front of it.
C. dongusensis had diagostic
rows of interlocking bony plates called osteoderms along their backs,
reaching from head to tail (Tverdokhlebova, 1972). These osteoderms
constitute the most frequently found fossilized remain of
Chroniosaurus. The flat osteoderm plates are connected to the neural
arches of vertebra by an extension of bone on their undersurfaces. Each
osteoderm thus is paired with a single vertebra. The front margin
of each osteoderm also has a pair of "anterior wings" that slip into a
notch in the posterior margin of the osteoderm in front of it.Fig.7: Skull of Chroniosaurus dongusensis (image: after Golubev 1998.)
Chroniosuchians are also distinguished from other early reptiliomorphs by the lack of intertemporal bones in the skull, as well as the presence of holes in front of the eye sockets, called antorbital fenestrae. Like many early tetrapods, chroniosuchians have vertebrae that are divided into three parts: a pleurocentrum and an intercentrum on the bottom, and a neural arch on top. Chroniosuchians have vertebrae whose pleurocentrum makes up most of the body, while the intercentrum is small and wedge-like.
.
References:
Benton, M.J. 2005. Vertebrate Paleontology. Blackwell Publishers.
Bratislava, Jozef Klembara 1995. The external gills and ornamentation of skull roof bones of the Lower Permian tetrapod Discosauriscus (Kuhn 1933) with remarks to its ontogeny. Paläontologische Zeitschrift. 69 (1): 265–281.
Brough, M.C. and Brough J. 1967. The Genus Gephyrostegus. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences no 252 (776): pp. 147–165.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution. W. H. Freeman & Co.
Carroll, R. L., 2002 Early land vertebrates. Nature, 418, pp 35-36
Clack, J.A, 2012. Gaining Ground. University of Indiana Press.
Gauthier, J., A.G. Kluge, and T. Rowe 1988. The early evolution of the Amniota. In The phylogeny and classification of the tetrapods, no 1: amphibians, reptiles, birds. Edited by M.J. Benton. Clarendon Press, Oxford, pp. 103-155.
Hildebrand, M. and G.E. Goslow 2001. Analysis of Vertebrate Structure. New York, John Wiley.
Holmes, R. 1984 The Carboniferous Amphibian Proterogyrinus scheelei Romer, and the Early Evolution of Tetrapods". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 306 (1130): 431–524.
Huxley, T.H. and E.P. Wright, 1867. On a collection of fossil vertebra from the Jarrow Colliery, County of Kilkenny, Ireland. Transactions of the Royal Irish Academy 24, pp. 51-368.
Jaeckel, O. 1902. Über Gephyrostegus bohemicus, n.g. n.sp. Zeitschrift der Deutschen Geologischen Gesellschaft no 54, pp. 127–132.
Kuhn, O. 1970, ed. Handbuch der Palaoherpetologie, Teil 5A. Fischer, Stuttgart.
Milner, A.R. 1980. he Tetrapod Assemblage in Nytany, Czechoslovakia. In Panchen, A.L. (ed.), The Territorial Environment and the Origin of Land Vertebrates. Systematics Assoc. Special Volume 15, Academic Press London, pp. 439-496.
Palaios 2014d, "Reptiliomorphs". Palaios.org website.
Panchen, A.L. 1970. Anthracosauria, in Kuhn, O. 1970, ed. Handbuch der Palaoherpetologie, Teil 5A. Fischer, Stuttgart.
Romer, A.S. 1928 . A Skeletal Model of the Primitive Reptile Seymouria, and the Phylogenetic Position of that type. The Journal of Geology. 36 (3): 248–260.
Romer, A.S. 1956. Vertebrate Paleontology. University of Chicago Press.
Romer, A.S. 1970. A new anthracosaurian labyrinthodont, Proterogyrinus scheelei, from the Lower Carboniferous. Kirtlandia. 10: 1–16.
Säve-Söderbergh, G. 1934. Some points of view concerning the evolution of the Vertebrates and the classification of this group. Arkiv för Zoologi 26A(17), pp. 1-20
Tatarinov, L. P., 1972, Paleontology and the phylogenetic laws of Lower Terrestrial Vertebrates. Palaeontological Journal, v.6, n.3, pp. 389-398.
Tverdokhlebova, G.I. 1972. A New Batrachosaur genus from the Upper Permian of the South Urals. Palaeontological Journal, v.6, n.1, pp. 95-103.
Tverdokhlebov, V.P., G. I. Tverdokhlebova, A. V. Minikh, M. V. Surkov, and M. J. Benton. 2005. Upper Permian vertebrates and their sedimentological context in the South Urals, Russia. Earth-Science Reviews 69, pp. 27-77
von Huene, F. 1956: Paläontologie und Phylogenie der niederen Tetrapoden, G. Fischer, Jena
Watson, D.M.S. 1918. On Seymouria, the most primitive known reptile. Proceedings of the Zoological Society of London. 88 (3–4): 267–301.
Williston, S.W. 1911a. A new family of reptiles from the Permian of New Mexico. American Journal of Science, Series 4, 31:378-398.
Williston, S.W. 1911b. Restoration of Seymouria baylorensis Broili, an American Cotylosaur. The Journal of Geology. 19 (3): 232–237.
Wright, E.P. and T.H. Huxley, 1866. On a collection of fossils from the Jarrow Colliery, County of Kilkenny, Ireland. Geol. Mag. (4), 3, pp. 165-171.
Glossary