Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
45. Homo erectus 2: Java
Discovery of the first Homo erectus specimen in the island of Java in southeast Asia was directly inspired by the 19th century “missing link” concept of a bipedal “ape-man,” who combined traits of today’s humans and apes. Both
 Charles Darwin and his contemporary Ernst Haeckel believed that upright
posture and bipedalism were direct precursors to tool making, by
freeing the hands.
Charles Darwin and his contemporary Ernst Haeckel believed that upright
posture and bipedalism were direct precursors to tool making, by
freeing the hands. Few if any Victorian evolutionists could have predicted that a gap of at least 3.5 million years separated the first evidence of bipedalism, now known from the Chad hominin Sahelanthropus dating 6 mya in the Late Miocene (Brunet et al. 2002), from the first stone tools at 2.5 mya. Meanwhile, however, the criteria of erect posture or bipedalism remains the most important morphological trait separating fossil hominids from apes. The evolutionary advantage of bipedalism most likely involved energy conservation in travelling. By the time of Homo erectus at ca. 1.8 mya, the human anatomy of long legs, tall stature, shorter arms, and full bipedalism had evolved (Wood and Collard 1999).
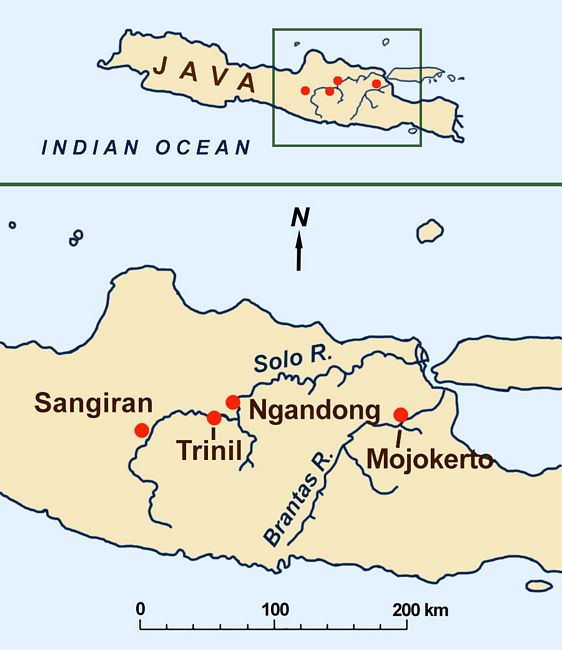
Fig.1: Map of major Homo erectus sites in east-central Java .
Eugene Dubois and the discovery of Homo erectus at Trinil
Eugene Dubois (1858-1940) was a Dutch doctor and anatomist who, in 1891, discovered the first fossils of Homo erectus in Trinil, Java. Indonesia (then a Dutch colony) was still only tentatively known to contain Pliocene and Pleistocene fossil beds, comparable to better documented zones in the Siwalik Hills of Pakistan. Discovery in 1878 of an extinct Pleistocene chimpanzee called Anthropithecus (“man-ape”) in the Siwalik beds
 attracted the attention of Dubois (fig.2). He became convinced of southern
Asia’s potential for “missing link”evidence, as did the unique presence
of the orangutan in Indonesia.
attracted the attention of Dubois (fig.2). He became convinced of southern
Asia’s potential for “missing link”evidence, as did the unique presence
of the orangutan in Indonesia.When Dubois began his fieldwork in the late 1880s, only Neanderthals had been identified as fossil hominids. Neanderthal remains from Germany and Belgium, in terms of skeletal and cranial form, appeared robust but basically modern (Huxley 1863). The quest to find a much earlier, more ape-like human ancestor was a revolutionary step initiated by Dubois.
Pleistocene fossil beds along the Solo River and Kendig Hills in east-central Java (named the Trinil or Kendig zone by Dubois) soon revealed faunal correspondences with the Siwalik Hills of Pakistan, including rhinocerous, hippopotami, a primitive elephant named Stegadon, buffalo, deer, hyenas, and large felines. In 1891, the Trinil beds produced a molar resembling Anthropithecus, and then the first remains of Homo erectus.
Fig.2: Eugene Dubois (photo: ca 1890).
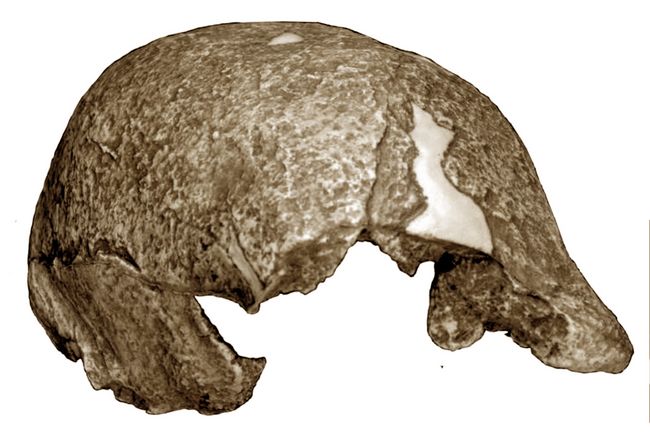
The 1891 finds included a thick skullcap or calvaria known as Trinil 2, with pronounced brow ridges and constriction behind the eye orbits (fig.3). Then, in 1892, Dubois found a human-like left femur (thighbone) which showed upright posture. This was at first grouped with the skullcap as a kind of advanced, bipedal chimpanzee named Anthropithecus erectus.
The Trinil 2 skullcap shows a low, sloping profile with a pinched-in, postorbital constriction, contrasting to the domelike form of the modern human skull. Thick walls and heavy brow ridges also appeared apelike. Further study
 of
the skullcap by Dubois, however, with removal of the
rock matrix in its cavity revealed a much larger cranial capacity (900
cc), twice that of a chimpanzee (ca. 450 cc). The Trinil 2 skullcap is
now dated by potassium-argon dating of its geological context to be
850,000-900,000 years old (fig.7).
of
the skullcap by Dubois, however, with removal of the
rock matrix in its cavity revealed a much larger cranial capacity (900
cc), twice that of a chimpanzee (ca. 450 cc). The Trinil 2 skullcap is
now dated by potassium-argon dating of its geological context to be
850,000-900,000 years old (fig.7).In a tribute to Haeckel, Dubois (1894) renamed the fossil Pithecanthropus erectus, “erect ape-man.” This taxon was to be eventually replaced (after discovery of the "Peking Man" fossils in the 1920s and 30s, and of the australopithecines in the 1920s-40s) by the name used today, Homo erectus (Mayr 1950).
Fig.3: Skullcap of Homo erectus from Trinil, Java (Yale Peabody Museum cast; photo: Athena Review).
Ngandong: Discovery of "Solo Man"
Two decades after the Trinil 2 discovery, 1909-10 German excavations led by Margarete Selenka across the Solo River from Trinil recovered Pleistocene fauna of a narrower and mainly later range than those found by Dubois. While the expedition found no hominid remains, it helped refine the Pleistocene biostratigraphy of Java.
Findings of the next fossil hominids in Java came in the early 1930s at Ngandong, on an upper terrace of the Solo River only ten km from Trinil (fig.1). In 1931-1933 the Dutch Geological Survey under W. F. F. Oppennoorth excavated rich Upper Pleistocene fossil deposits some 20 -24 m above the current level of the Solo. Besides thousands of animal fossils (extinct buffalo, wild oxen, Stegodon, pigs, tigers, etc.), the project recovered 12 hominid calvaria, the largest sample from a single Javan site.
 The
Ngandong remains, named Homo soloensis by Oppennoorth (1932), and now classed as Homo erectus soloensis, represent
a much-debated group of Upper Pleistocene hominids. They had
significantly larger cranial capacities than earlier Javan Homo
erectus, averaging 1210 cc compared to 883 cc (Jacob 1981). Massive
brow ridges (fig.4) and thick cranial bones, however, make “Solo Man”
(dated 120,000-110,000 BP) appear still closely linked to Homo
erectus.
The
Ngandong remains, named Homo soloensis by Oppennoorth (1932), and now classed as Homo erectus soloensis, represent
a much-debated group of Upper Pleistocene hominids. They had
significantly larger cranial capacities than earlier Javan Homo
erectus, averaging 1210 cc compared to 883 cc (Jacob 1981). Massive
brow ridges (fig.4) and thick cranial bones, however, make “Solo Man”
(dated 120,000-110,000 BP) appear still closely linked to Homo
erectus.Fig.4: Solo 6 skull from Ngandong, Java (photo: Athena Review, from cast at AMNH).
Soon afterwards, in the mid-to-late1930s, paleontologist Ralph von Koenigswald identified an important series of Lower and Middle Pleistocene Homo erectus fossils collected by Javan farmers. Originally working with the Dutch Geological Survey to classify fossil fauna, von Koenigswald obtained funding from the Carnegie Institute of Washington DC for early hominid site exploration. In the course of these 1930s projects, detailed geological maps were made in Java (Shipman 2001; Huffman 2001).
Sangiran and Mojokerto
At Sangiran, one of several large volcanic domes in east-central Java (which ca. 1.5 mya formed a lake shore and riverine zone), von Koenigswald found a Homo erectus mandible in 1934. Then, in 1937, he discovered a skullcap called Sangiran 2, similar to Trinil 2 but with a smaller cranial capacity of 815 cc; and, in 1938-9, several early Homo erectus skulls and mandibles with massive teeth. One fossil, Sangiran 6, was at first classed as Paranthropus (or Australopithecus) robustus. Another, Sangiran 4, has an australopithecine-like diastema or gap between the upper canines and incisors. Both are now considered to predate 1.5 mya.
 The
fossil cranium known as Sangiran 17 (fig.5) was later discovered at Sangiran in 1969 by Towikromo. The most
complete Homo erectus skull found in East Asia, it dates from 1.15 mya
years ago (fig.7). It has the prominent brow ridges and thick skull bones
typical of Homo erectus, and a relatively large brain size. The cranial
capacity is estimated at 1004 cc, equivalent to those of some Homo
erectus crania found in China, and slightly larger than that found at
Trinil in east Java (Wu et al 2010)
The
fossil cranium known as Sangiran 17 (fig.5) was later discovered at Sangiran in 1969 by Towikromo. The most
complete Homo erectus skull found in East Asia, it dates from 1.15 mya
years ago (fig.7). It has the prominent brow ridges and thick skull bones
typical of Homo erectus, and a relatively large brain size. The cranial
capacity is estimated at 1004 cc, equivalent to those of some Homo
erectus crania found in China, and slightly larger than that found at
Trinil in east Java (Wu et al 2010)Fig.5: The Sangiran 17 skull, dated at about 1.15 mya (photo: Athena Review, from cast at AMNH).
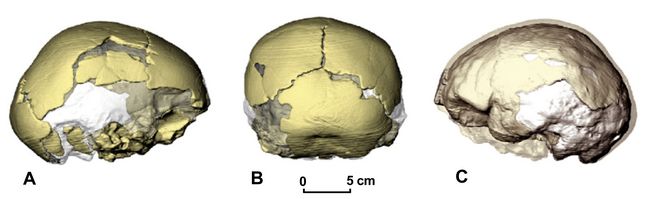
An even earlier Homo erectus fossil of a child’s skull was recovered by von Koenigswald in 1936 in the east Javan village of Perning near Mojokerto, 180 km east of Sangiran on the Brantus River. Now dated from 1.55 mya to as early as 1.8 mya, the Perning skull came from a layer representing an ancient marine deltaic setting. A brain endocast has been measured from the skull, showing a cranial capacity of 620-640 cc. for what has been identified as a four-year-old child (Balzeau et al. 2005) .
The fossils from Sangiran and Mojokerto demonstrate that initial hominid migrations to southeast Asia entered a diverse range of local environments.
Fig.6: Mojokerto child's skull from Perning, viewed from A) right side; B) rear, occipital view; and c) left side, showing brain endocast (after Balzeau et al. 2005).
In 1939, Von Koenigswald took a number of Javan Homo erectus fossils to China to compare with the Zhoukoudian hominid remains being studied by Franz Weidenreich. Similarities seen by von Koenigswald and Weidenreich (1939) eventually led to combined use of the Homo erectus taxon for both “Java Man” and “Peking Man” (Mayr 1950).
During World War II, von Koenigswald managed to safeguard many of the Javan fossils even though he was imprisoned by the Japanese from 1942-5. From the 1960s-80s, Tekeu Jacob directed numerous paleoanthropological projects in Java, including excavations at Ngandong, Sambungmachan, and Sangiran revealing several more Homo erectus and H. soloensis skulls (Jacob 1973, 1981). Since the 1990s, work by international teams has progressed at a number of sites including Mojokerto, Ngandong, and Sangiran Dome. Especially interesting finds are Oldowan stone tools and faunal remains with butchery marks recovered in the Bapang Formation in the Ngebung Hills (Semah et al. 1992).
Dating Homo erectus sites in Java
Before the advent of potassium-based geochronology, the dating of fossil deposits in Java was done by biostratigraphy, associating mammalian taxa with geological layers. In 1969, geochronologist Garniss Curtis first applied potassium-argon dating to a volcanic rock from the Mojokerto site, producing a date of 1.9 mya (Swisher et al. 2000). This was basically upheld over 20 years later by more accurate Ar/Ar dates from 1.81-1.6 mya on pumice from the Pucangan (Sangiran) Formation (Swisher et al. 1994). An ind
 ependent series of dates obtained for the overlying Kabuh
(Bapang) Formation in central Java shows a group of five Homo erectus
fossils predating 1.51 mya (Larick et al. 2001; Larick et al. 2004).
ependent series of dates obtained for the overlying Kabuh
(Bapang) Formation in central Java shows a group of five Homo erectus
fossils predating 1.51 mya (Larick et al. 2001; Larick et al. 2004).These early dates remain controversial. If the oldest dates are accurate, correlations with paleomagnetic stratigraphy from the Rift Valley of Africa would show the arrival of hominids in Java as early as the Olduvai subchron (1.98-1.79 mya). Significantly, such early dates suggest the African emergence of Homo and the initial dispersal to subtropical Asia may be directly linked. Conditions favoring this initial migration center on the geologically active Tethys Corridor, a tectonic zone along the south edge of the European continental plate. Environmental change created new subsistence niches at the start of the Pleistocene, while lowered sea-levels due to glaciation exposed the Sunda shelf, over which hominids could walk to southeast Asia.
Fig.7: Time chart of Homo erectus finds in Java, correlated with Pleistocene stages (after Larick et al 2004).
Based on this recent dating, the earliest Homo erectus groups settled in Java by 1.8-1.6 mya around coastal deltaic swamps and lakes at the south end of the Sunda shelf, associated with the Sangiran formation and the Ci Saat fauna (including the large carnivore Panthera and bovids such as deer and hippopotami). Homo erectus fossils from this oldest level include the primitive-looking Sangiran 4 found by von Koenigswald, as well as four other individuals (Sangiran 27, 31, 41, and 57) found since 1978.
At about 1.5 mya the Sangiran levels were superimposed by river and lake sediments of the Kabuh Formation with associated Trinil Fauna (Dubois’ original Trinil zone). Homo erectus finds include the relatively complete Sangiran 17 cranium (fig.5) found in 1969; the Sangiran 50 cranium and maxilla found in 1993 at Tanjung (see Larick et al. 2004); and the Sangiran 2 skullcap of von Koenigswald (fig.8), all three dated at 1.25-1.0 mya, slightly earlier than the Trinil 2 calotte.
 After
800,000 BP, fossil evidence ceases at the Sangiran Dome. Around this
time Homo erectus may have rafted across 25 km of ocean to the island
of Flores, east of Bali, where Oldowan tools have been found (Morwood
et al. 1998). More recent finds by Morwood and colleagues on Flores
show that very late, small Homo erectus-like hominids named Homo
floresiensis still lived there from 95,000-13,000 BP, overlapping the
time frame of "Solo Man," and even living into the period of early
animal and plant domestication by modern humans in the Near East.
After
800,000 BP, fossil evidence ceases at the Sangiran Dome. Around this
time Homo erectus may have rafted across 25 km of ocean to the island
of Flores, east of Bali, where Oldowan tools have been found (Morwood
et al. 1998). More recent finds by Morwood and colleagues on Flores
show that very late, small Homo erectus-like hominids named Homo
floresiensis still lived there from 95,000-13,000 BP, overlapping the
time frame of "Solo Man," and even living into the period of early
animal and plant domestication by modern humans in the Near East.Fig.8: Homo erectus Sangiran 2 skull cap, dated at about 1 million years ago (U.S. National Museum cast).
After a hiatus of about 600,000 years after Trinil, the Homo soloensis remains found in the Upper Pleistocene Notopuro and Ngandong Formations represent a much more recent fossil hominid group from Java.. Uranium-series and electron spin resonance (ESR) dates of 53-27,000 BP have been obtained on bovid teeth from the same Ngandang deposit producing the Homo erectus soloensis skulls (Swisher et al.1997). These dates plus anatomical evidence provide grounds to identify the Ngandong fossils as part of a very late Homo erectus population contemporary with (and a separate species from) Homo sapiens (Antón 2002). The Ngandong fossils remain an important enigma, partly contemporaneous with both the newly found Homo floresiensis of the Indonesian island Flores, and the relatively large-brained fossil skulls from Dali and Mapa in China, representing late Asiatic descendants of Homo erectus.
References:
Antón, S.C., 2002. “Evolutionary significance of cranial variation in Asian Homo erectus.” Amer. Journal of Physical Anthropology 118: 301-323.
Balzeau, A., D. Grimaud-Herve, and J. Teuku 2005. Internal cranial features of the Mojokerto child fossil (East Java, Indonesia). Journal of Human Evolution 48 535-553.
Brunet, M., et al. 2002. “A new Hominid from the Upper Miocene of Chad, Central Africa.” Nature 418:145-151.
Dubois, Eugene. 1894. Pithecanthropus erectus. Eine menschenaehnliche Uebergangsform aus Java. Batavia, Landsdrukkerij.
Dubois, Eugene. 1896. “On Pithecanthropus erectus, a transitional form between man and the apes.” Trans.Royal Dublin Soc., ser.2, vol.6, pp. 1-18.
Haeckel, E. 1892 (orig.1868). The History of Creation. (4th Eng. ed.) London, Kegan Paul, Trench, Trubner & Co.
Huffman, O.F. 2001. “Geologic context and age of the Perning/Mojokerto Homo erectus, East Java.” J.of Human Evolution 40:353-62.
Huxley, T. H. 1863. “On Some Fossil Remains of Man,” in Evidence as to Man’s Place in Nature. London, Williams and Norgate.
Jacob, T. 1973. “Palaoanthropological discoveries in Indonesia with special reference to the finds of the last two decades.” J. of Human Evolution 2:473-85.
Jacob, T. 1981. “Solo Man and Peking Man.” in B.A. Sigmon & J.S. Cybulski (eds.), op. cit., pp.87-104.
Larick, R. and R.L. Ciochon 1996. “The African Emergence and dispersal of the genus Homo.” American Scientist 84:538-552.
Larick, R. et al. 2001. “Early Pleistocene 40Ar/39/Ar ages for Bapang Formation hominins, Central Java, Indonesia.” Proc. Nat. Acad. of Sciences 98:4866-71.
Mayr, E. 1950. “Taxonomic categories in fossil hominids.” Cold Spring Harbor Symposia on Quantitative Biology 15:109-118.
Morwood, M. et al. 1998. “Fission-track ages of stone tools and fossils on the east Indonesian island of Flores.” Nature 392: 173-176.
Oppennoorth, W.E.F. 1932. “Homo (Javanthropus) soloensis: Een pleistocene mensch van Java.” Wetenschappelijke medeligen Dienst van den Mijnbrouw in Nederlandsch-Indië 20:49-63.
Shipman, P. 2001. The Man Who Found the Missing Link. New York, Simon & Schuster.
Swisher, CC., G.H. Curtis, Y. Jacob, A.G. Getty, A. Suprojo, Widiasmoro. 1994. “Age of the earliest known hominids in Java, Indonesia. Science 263:1118-1121.
Swisher C.C., W.J. Rink, H.P. Schwarcz, and S.C. Antón 1997. “Dating the Ngandong Humans.” Science 276: 1575-1576.
Swisher, C.C., G.H. Curtis, and R. Lewin. 2000. Java Man. New York, Scribner.
Von Koenigswald, G.H.R. 1956. Meeting Prehistoric Man. Thames and Hudson.
Von Koenigswald, G.H.R and F. Weidenreich 1939. “The relationship between Pithecanthropus and Sinanthropus.” Nature 144: 926-929
Wood, B. and M. Collard 1999. “The Human Genus.” Science 284:65-71.
Glossary