Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
39. New World Monkeys: Late Eocene to Miocene
Platyrrhine Monkeys
New World Monkeys are called Platyrrhini ("flat-nosed"), in contrast to their Old World cousins, the Catyrrhinii ("sharp-nosed"). Current evidence shows that New World monkeys (Platyrrhini) first appeared in South America in the Late Eocene period, about 36 million years ago. Numerous similarites in the anatomy of both fossil and extant New World Monkeys with Late Eocene and Early Oligocene Old World anthropoid fossil taxa, such as Proteopithecus from the Late Eocene Fayum in Egypt (Simons 1997), provide strong evidence that New World Monkeys originally came from Africa.
South America was an island continent geographically isolated from Africa as a result of plate tectonics or continental drift more than 65 million years ago. Based on similarities with African taxa, it is evident that the first New World Monkeys, in some manner, rafted across the Atlantic Ocean to South America during the Late Eocene or Early Oligocene. Alternative fossil evidence that monkeys migrated by land from Africa or Asia to the Americas via the Bering Strait is currenntly scanty or non-existent. The problem of how early Old World monkeys reached the New World thus remains unresolved (cf. Ciochon ed. 1984)
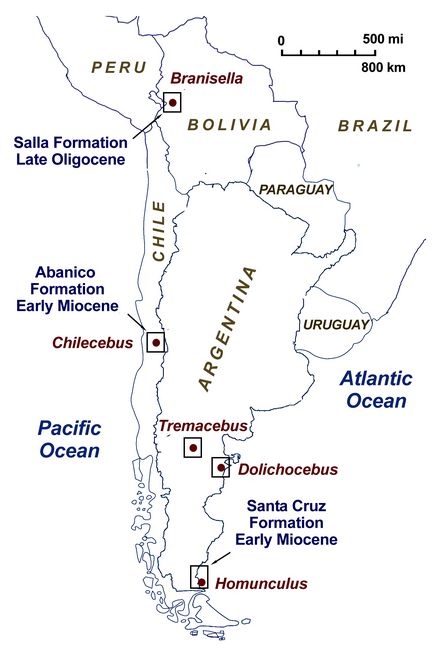
In 1840, the first fossil of a New World Monkey was found in Brazil, from a Pleistocene platyrrhine called Protopithecus brasiliensis (Lund 1840). In 2015, findings of a Late Eocene primate were reported, for Perupithecus, dated at 36 mya (Bond et al. 2015). Before then, the earliest fossil evidence of New World Monkeys (Platyrrhines) was the late Oligocene Branisella, discovered in Bolivia in the 1960s (fig.1). Currently, besides numerous teeth and jaw fragments from dozens of fossil taxa, five crania of fossil New World platyrrhines are known, belonging to Tremacebus, Dolichocebus, Chilecebus, Homunculus, and Lagonimico. These fossils were found in Argentina and Chile (fig.1).
Fig.1: Map showing locations of several New World Monkey fossil sites in South America.
Perupithecus
Fossils from three early Platyrrhine monkeys in Peru dating from about 36 mya were discovered by paleontologist Kenneth Campbell and his associates in 2010 (Bond et al. 2015). The characteristics of the teeth of these early monkeys provide the first evidence that South American monkeys actually have an African ancestry.The Perupithecus tooth and jaw fossils show features that suggest links to African anthropoids of similar age.
The Perupithecus fossils have pushed back the dates of the earliest platyrrine monkeys by 10 million years. They indicate the importance tropical forest in the evolution of South American primates. an environment distinct from that of higher-latitude primates known from Bolivia, Argentina, and Chile from the Late Oligocene and Early Miocene, including Branisella, Szalatavus, Tremacebus, Dolichobebus, and Chilecebus, all of whom date in the range of 26-16 mya.
Branisella
Branisella boliviana is from the Salla Formation of Bolivia, dating from 26 mya (Rosenberger et al. 1991). It was discovered in the early 1960s by the paleontologist Leonardo Branisa, and named after him by Hoffstetter (1969), who who first described and classified the fossil remains. These consist of jaw fragments and teeth (fig.2). For many years it was the earliest known fossil of a Platyrrhine monkey.

In 1996 a pair of upper and lower jaw fragments and another nearly complete mandible of Branisella were recovered (Takai et al. 2000). These new fossil materials reveal several morphological features in Branisella: Regarding premolars, P2 is much smaller than P3,4, whereas P2 is relatively small but probably sexually dimorphic. Secondly, the zygomatic arch resembles those in extant Callicebus (titi monkey). Thirdly, the mandibular arcade is nearly V-shaped and is distinct in form from those of both callitrichines (marmosets and tamarins) and Cebus (capuchin monkey). Fourthly, the upper and lower molars wore down rapidly in life, suggesting a herbivorous diet and the possibility of terrestriality. Finally, the dental eruptive sequence is the same as in extant Aotus (owl monkey) (Takai et al. 2000).
Fig.2: Upper and lower jaw fossils of Branisella boliviana
Branisella boliviana had a dental formula of 2:1:3:3 on both the upper and lower jaws. The molars have low, rounded cusps, and the upper molars possess a hypocone and are quadritubercular. Based on the mandible and the small premolar (P2), this species had a short face. It probably had an average body mass of about 1.0 kilograms (Fleagle, 1988; Conroy 1990). Within platyrrhines, this taxon has been interpreted as either a stem platyrrhine not related to any of the living forms or as a primitive callitrichine.
Morphologically, Branisella is similar to Proteopithecus (Simons 1997), a late Eocene primate from the Fayum region of Africa, in its reduced upper 2nd premolar and unreduced lower 2nd premolar, although not in the lower canine morphology. This suggests a close phyletic relationship between them, evidence that the primitive Platyrrhine ancestors of Branisella likely came to South America from Africa (Takai et al. 2000).
Tremacebus
Tremacebus harringtoni was a platyrrhine monkey in the family Cebidae and the subfamily Aotina.who lived in Argentina in the early Miocene, about 20 mya (Rosenberger 1985; Fleagle 1988). It is thought to be an ancestor of the extant genus Aotus.
 T. harringtoni
is noted for its orbital enlargement (Hershkovitz,
1974), a feature that led Rosenberger (1979, 1984) and Szalay and
Delson (1979) to conclude that it has phylogenetic affinities with Aotus, and that nocturnality in the Aotus clade extends back at least to the early Miocene. However,
though relative orbit size in Tremacebus falls within the range of
nocturnal strepsirrhines, it is relatively smaller than the orbits of
nocturnal haplorhines Aotus and Tarsius (Kay and Kirk, 2000).
T. harringtoni
is noted for its orbital enlargement (Hershkovitz,
1974), a feature that led Rosenberger (1979, 1984) and Szalay and
Delson (1979) to conclude that it has phylogenetic affinities with Aotus, and that nocturnality in the Aotus clade extends back at least to the early Miocene. However,
though relative orbit size in Tremacebus falls within the range of
nocturnal strepsirrhines, it is relatively smaller than the orbits of
nocturnal haplorhines Aotus and Tarsius (Kay and Kirk, 2000).Aotus also has the largest relative olfactory fossa breadth and the largest olfactory bulb volume compared to brain volume among , and relies more on olfactory cues when foraging. Tremacebus had a much smaller olfactory fossa breadth than the noctural Aotus. This suggests that perhaps this species was more diurnal in behavior (Hershkovitz 1974).
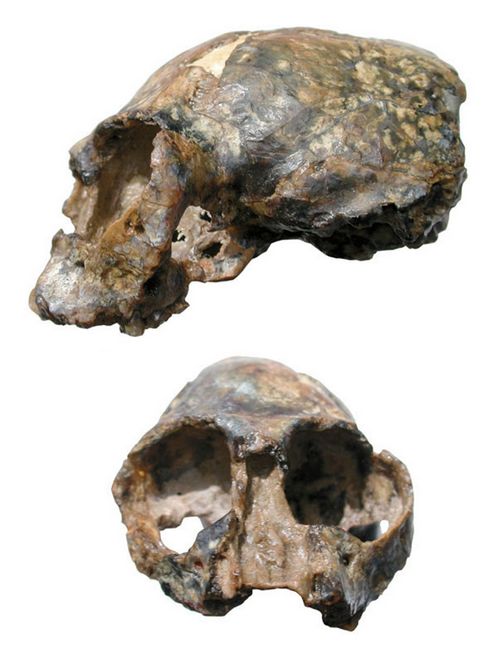
Fig.3: Skull of Tremacebus harringtoni (after Kay et al.2004)
T. harringtoni had a short, broad snout and orbits between extant diurnal platyrrhines and extant members of the genus Aotus in size. The skull (fig.3) had well defined temporal lines (Conroy, 1990), and an auditory bulla similar to that found in extant platyrrhines (Martin 1990). Its average body mass was about 1.8 kilograms (Fleagle 1988).
T. harringtoni had a dental formula of 2:1:3:3 on the upper jaw.The dentition of T. harringoni includes quadrate upper molars with a broad lingual cingulum. Based upon its dental morphology, this was most likely an herbivore (Conroy 1990).
Dolichocebus
 Dolichocebus was a New World Monkey in the Cebidae family who lived in Argentina during the Early Miocene. The type species is D. gaimanensis, found in the Trelew Member of the Sarmiento Formation near the village of Gaiman, Chubut Province, Argentina, dated to between 21.5-17.0 mya (Fleagle and Kay 1989).
Dolichocebus was a New World Monkey in the Cebidae family who lived in Argentina during the Early Miocene. The type species is D. gaimanensis, found in the Trelew Member of the Sarmiento Formation near the village of Gaiman, Chubut Province, Argentina, dated to between 21.5-17.0 mya (Fleagle and Kay 1989). Dolichocebus is known from the type skull encased in a concretion, as well as numerous isolated teeth, parts of two mandibles, and a talus. Analysis indicatess that Dolichocebus is a stem platyrrhine, as are late Oligocene Branisella and early Miocene Tremacebus, Soriacebus, and Carlocebus.
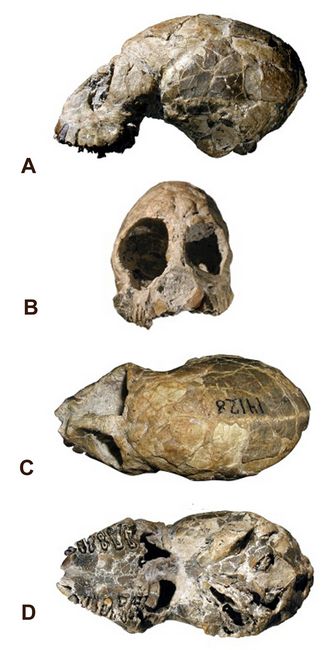
Kay et al. (2008) analyzed all Dolichocebus fossil material using conventional surface anatomy and micro-CT data from the cranium (fig.4). Newly imaged internal anatomy of the skull demonstrated that anatomical characters hitherto supposed to support a phyletic link between Dolichocebus and either callitrichines (marmosets, tamarins, and Callimico) or Saimiri (squirrel monkeys) are either indeterminate or absent.
Fig.4: Skull of Dolichocebus gaimanensis, with four views: A) lateral; B) frontal; C) top (dorsal); D:) bottom (ventral) (after Kay et al.2008).
A review of all available phylogenetic data suggests a layered evolutionary pattern, with several independent extinct clades leading to modern platyrrhine groupings over a nine-million-year interval in the Miocene. Homoplasy or convergence in dental and cranial characters is seen as present at all levels of biological analysis of New World Monkeys, from amino-acid sequences to aspects of adult bony morphology, behavior, and adaptation (Kay et al. 2008).
Chilecebus
Chilecebus carrascoensis was a small Platyrrhine monkey that lived in Chile during the Early Miocene period. It is a member of the family Cebidae.
A nearly complete skull of C. carrascoenis was found at Rio las Lenas, in the Abanico Formation in the Andes of central Chile (Flynn et al. 1995). An argon/Argon (40Ar/39Ar) date taken from rocks associated with the skull indicates an Early Miocene age of ca. 20 mya.
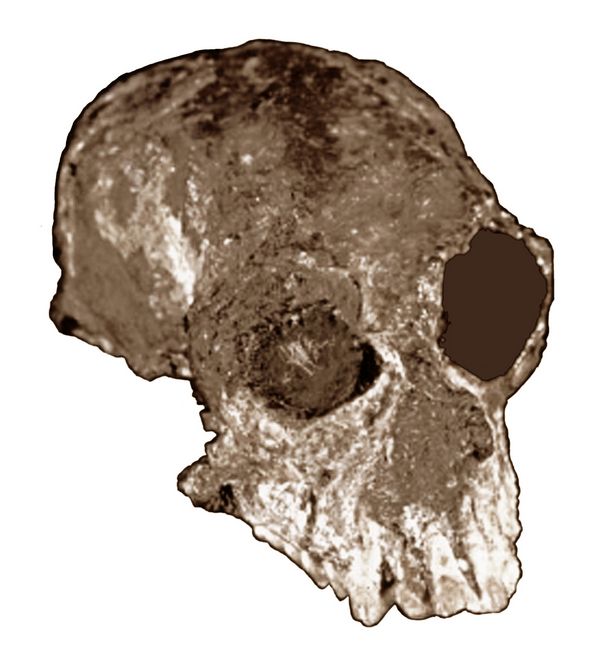
The skull (fig.5), only about 3 cm tall, retains both eye sockets and all teeth in the upper jaw. Its dentition indicates that C. carrascoensis was a fruit or leaf eater. Chilecebus had a body mass of about 1.3 pounds (600 grams).
Flynn and his coworkers at AMNH studied the tiny skull and its brain cavity using CAT scans, determining that Chilecebus provided evidence that increases in anthropoid brain sizes must have occurred independently several times. A small olfactory bulb in Chilecebus was not counterbalanced by an amplified visual system. This finding indicates that in primate evolution the visual and olfactory systems were not as tightly coupled as widely assumed. In many modern primates, the size of the visual and olfactory centers in the brain are negatively correlated, with visually acute primates typically having weaker senses of smell (AMNH 2019).
Fig.5: Skull of Chilecebus carrascoensis (after Flynn et al. 1995)
Chilecebus is thought to be diunal, based on the size of the opening for the optic nerve. The infolding (sulcus) pattern of its brain surface, while simpler than in most modern anthropoids, is relatively complex for such an ancient primate, as it possesses at least seven pairs of sulcal grooves.
.
Homunculus
Homunculus patagonicus was a ceboid monkey who lived in Argentina during the Miocene. It was discovered in 1891 by Carlos Ameghino in the southeast of Santa Cruz Formation. It is likely the ancestral species for the subfamily Pitheciinae, whose living members include the sakis, bearded sakis, and the uakaris.
Homunculus patagonicus only was the second fossil platyrrhine discovered in South America. Between 1891 and 1894, Ameghino found mandibular fragments, a partial cranium, and postcranial e
 lements. The holotype of H. patagonicus (MACN-A
634) was collected by Ameghino from the north shore of the estuary of
the Rio Gallegos. Most of the holotype collection was unfortunately
later lost.
lements. The holotype of H. patagonicus (MACN-A
634) was collected by Ameghino from the north shore of the estuary of
the Rio Gallegos. Most of the holotype collection was unfortunately
later lost.A neotype, MACN-A 5757, was the best preserved mandible from the original Ameghino collection, likely discovered in the same general area as Ameghino’s type. Its morphology strongly resembles that of modern pitheciines and their fossil relatives but tends to be more primitive (Tejedor and Rosenberg 2008), .
Fig.6: H. patagonicus cranium, CORD-PZ 1130 from Puerto Estancia la Costa (Tauber 1991; after Perry et al. 2014)
In 2007 Perry et al. (2014) found a juvenile cranial monkey specimen of H. patagonicus at Puerto Estancia La Costa (Perry et al., 2010; Kay et al., 2012). The new material preserves unworn upper molars and undistorted facial anatomy, and improved the definition on extant H. patagonicus fossils, including a crania (CORD PZ-1130) (fig.6) preserving most of the left side, found at the same site (Tauber 1991). These specimens date from about 17.5 mya.
Orbital convergence in the juvenile cranium of H patagonicus resembles that of callitrichine platyrrhines, which have less convergence than extant non-callitrichine platyrrhine. Such orbital convergence also occurs in Tremacebus harringtoni, Dolichocebus gaimanensis, and in the Homuculus skull CORD-PZ 1130 (Hershkovitz, 1974; Kay et al., 2008, 2012)
Like these other forms, Homunculus presents a derived incisor-canine complex, a combination of features that parallels some of the advanced callitrichines, Callithrix and Cebuella. The mandibular profile is remarkably similar to Aotus. The partial cranium from the original series is notable for its flat frontal trigon, wide interorbital region, unenlarged orbits, deep maxilla and large maxillary sinus, and premaxilla.
Long bones preserve limb proportions similar to those of modern quadrupedal platyrrhines, such as Aotus and Callicebus, which lack advanced leaping adaptations. Overall, Homunculus appears to have been a primitive pitheciine, with versatile adaptations resembling the quadrupedal, diurnal titi monkeys, including an emphasis on feeding on tough fruits. it was likely a diurnal, foliovore-frugivore that ingested foods containing silica phytoliths or quantities of grit (Tejedor and Rosenberg 2008)
Cebupithecia
 Cebupithecia was a
New World monkey in the cebidae family found in Colombia, dating
from the Middle Miocene, Laventan stage (13.8 to 11.8 mya). The type species is C. sarmientoi. An incomplete skeleton and skull was discovered in 1944-45 at the Konzentrat-Lagerstätte of La Venta, part of the Honda Group in Colombia (fig.8), dated at about 13.5 mya (Striton 1951).
Cebupithecia was a
New World monkey in the cebidae family found in Colombia, dating
from the Middle Miocene, Laventan stage (13.8 to 11.8 mya). The type species is C. sarmientoi. An incomplete skeleton and skull was discovered in 1944-45 at the Konzentrat-Lagerstätte of La Venta, part of the Honda Group in Colombia (fig.8), dated at about 13.5 mya (Striton 1951).Cebupithecia sarmientoi had a dental formula of 2:1:3:3 on the lower jaw. The incisors of this species were procumbant and the canines had a stout appearance, while the molars were relatively flat (Fleagle, 1988). On the skull base, the ectotympanic ring is fused to the auditory bulla.
Fig.7: Skull of Cebupithecia.
Based upon its dental morphology, Cebupithecia ate mainly fruit, while seeds also played an important part in the diet. Cebupithecia sarmientoi had an average body mass of around 2.2 kilograms. Based on the postcranial remains, C. sarmientoi most likely relied more on vertical clinging and leaping than on quadrupedal locomotion, much like members of the extant genus Pithecia (Fleagle, 1988).The axial skeleton of C. sarmientoi display features associated with adaptations for clinging and leaping as in that genus. Cebupit
 hecia has a relatively long non-prehensile tail, which is most similar in absolute dimensions and proportions to that of Pithecia monachus.
hecia has a relatively long non-prehensile tail, which is most similar in absolute dimensions and proportions to that of Pithecia monachus.The Honda Group, and more precisely the co-called "Monkey Beds", are one of the most productive sites for fossil primates in South America (fig.8). Monkeys of the Honda Group may have been living in a habitat in contact with the Amazon and Orinoco Basins, while the La Venta area itself was probably a seasonally dry forest. Other fossil monkeys discovered in the same level where Cebupithecia was been found include Aotus dindensis, Micodon, Mohanamico, Saimiri annectens, Saimiri fieldsi and Stirtonia tatacoensis.
Fig.8: Location of the La Venta fossil beds in the Hondo Group of Colombia.
References:
Ameghino, F. 1891. Los monos fósiles del Eoceno de la República Argentina. Rev. Arg. de Hist. Nat. (Buenos
Aires) 1, 383–397
American Museum of Natural History (AMNH) 2019. 20-Million-Year-Old Skull Reveals Anthropoid Primate Brains Evolved More Than Once. AMNH.org website.
Bond, M., M.F.Tejedor, K.E.. Campbell Jr., L. Chornogubsky, N. Novo, and F.Goin. 2015. Eocene primates of South America and the African origins of New World. Nature 520. 538–546.
Conroy, G.C. 1990. Primate Evolution. W.W. Norton and Co.: New York.
Defler, T. 2018 Platyrrhine Monkeys: The Fossil Evidence, in History of Terrestrial Mammals in South America, pp. 161-184
Fleagle, J.G. 1988. Primate Adaptation and Evolution. Academic Press: New York.
Fleagle, J.G. and R.F. Kay 1989. The dental morphology of Dolichocebus gaimanensis, a fossil monkey from Argentina. American Journal of Physical Anthropology. 78 (2)
Flynn, J.J., A.R. Wyss, R. Charrier, C.C. Swisher et al. 1995. An Early Miocene anthropoid skull from the Chilean Andes. Nature 373, 603 - 607
Hershkovitz, P. 1970. Notes on Tertiary Platyrrhine Monkeys and Description of a New Genus from the Late Miocene of Colombia. Folia Primatol., Vol. 12, 1-37.
Hershkovitz, P. 1974. A New Genus of Late Oligocene Monkey (Cebidae, Platyrrhine) with Notes on Post Orbital Closure and Platyrrhine Evolution. Folia Primatol., Vol. 21, 1-35.
Hoffstetter M.R. 1969. Un primate de l'Pliocène inférieur sudamericain: Branisella boliviana gen. et sp. nov.. C. R. Acad. Sci. Paris 269: 434–437.
Kay, R.F., V.M. Campbell, J.BN. Rossie, M.W. Colbert, and T.B. Rowe 2004. Olfactory Fossa of Tremacebus harringtoni (Platyrrhini, Early Miocene, Sacanana, Argentina): Implications for Activity Pattern. The Anatomical Record, Part A. 281A: 1157–1172 .
Kay, R.F., J.G.Fleagle, T.R.T.Mitchell, M.W. Colbert, T. Bown, and D. W.Powers 2008. The anatomy of Dolichocebus gaimanensis, a stem platyrrhine monkey from Argentina Jour. Human Evol. 54,(3) 23-382
Lund, P. 1840. Nouvelles recherches sur la faune fossile du Bresil. Ann. Sci. Nat. (Paris) 13, 310–319
Martin, R.D. 1990. Primate Origins and Evolution: A Phylogenetic Reconstruction. Princeton University Press: Princeton, New Jersey.
Organ, Jason M., and Pierre Lemelin. 2011. Tail Architecture and Function of Cebupithecia sarmientoi, a Middle Miocene Platyrrhine from La Venta, Colombia. The Anatomical Record 294.12. 2013–2023.
Perry, J. M.G., R.F. Kay, S.F. Vizcaíno, and M.S.Bargo 2014. Oldest known cranium of a juvenile New World monkey (Early Miocene, Patagonia, Argentina): Implications for the taxonomy and the molar eruption pattern of early platyrrhines. Journal of Human Evolution. 74: 67–81. ,
Rosenberger, A.L. 1984. Fossil New World Monkeys Dispute the Molecular Clock. Journal of Human Evolution, Vol. 13, 737-742.
Rosenberger, A.L., W.C. Hartwig, and R.G. Woolf, 1991. Szalatavus attricuspis, an Early Platyrrhine primate. Folia Primatol. 56: 225-233.
Simons, E.L. 1997. Preliminary description of the cranium of Proteopithecus sylviae, an Egyptian late Eocene anthropoidean primate. Proc.Nat.Acad.Sci. no. 94, pp.14970-14975.
Stirton, R.G. 1951. Ceboid Monkeys from the Miocene of Colombia. Univ. Calif. Publs, Bull. Dep. Geol. Sci., Vol. 28, 315-356.
Takai, M F., S. Anaya, Nobuo Shigehara, and Takeshi Setoguchi 2000. New fossil materials of the earliest new world monkey, Branisella boliviana, and the problem of platyrrhine origins. Am J Phys Anthropol 111:263-281
Tejedor, Marcelo F.; Rosenberger, Alfred L. (2008). "A neotype for Homunculus patagonicus Ameghino, 1891, and a new interpretation of the taxon" PaleoAnthropology: 68–82.
Tejedor, Marcelo F.; Adán A. Tauber; Alfred L. Rosenberger; Carl C. Swisher III; María E. Palacios (2006-03-27). "New primate genus from the Miocene of Argentina". Proceedings of the National Academy of Sciences. 103 (14): 5437–41.
Glossary