Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
35. Early Primates 3
Anthropoidea
.
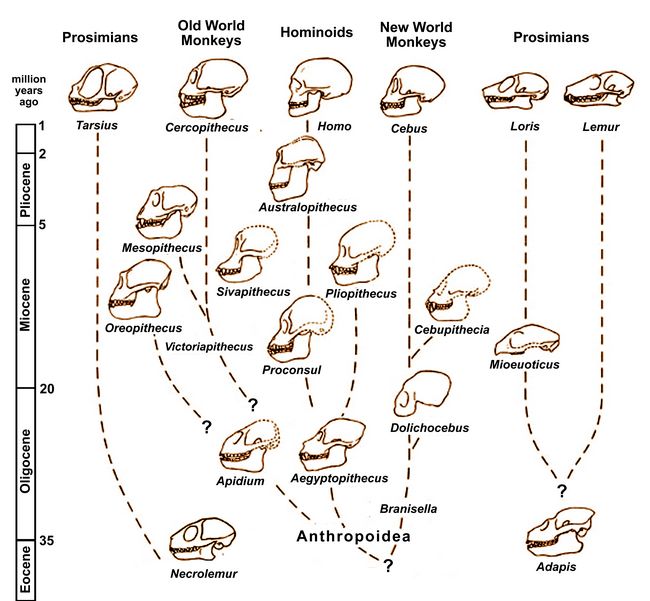 Anthropoidea, a suborder of primates, includes New World Monkeys,
Old World Monkeys, and Hominoidea (Apes and Humans). Major
trends of the evolutionary sequence of anthropoids from the late
Eocene to present are summarized in fig.1. By the Late Eocene
period (45-35 mya), Prosimians (Tarsiers, Lorises, and Lemurs) had
split off from Anthropoidea as a distinct suborder.
Anthropoidea, a suborder of primates, includes New World Monkeys,
Old World Monkeys, and Hominoidea (Apes and Humans). Major
trends of the evolutionary sequence of anthropoids from the late
Eocene to present are summarized in fig.1. By the Late Eocene
period (45-35 mya), Prosimians (Tarsiers, Lorises, and Lemurs) had
split off from Anthropoidea as a distinct suborder.Fig.1: Chronological chart showing relations between developing Anthropoid taxa (after Gingerich 1984).
During the Oligocene period (35-20 mya), within the Anthropoidea, the ancestors of Old World Monkeys and New World Monkeys split off from the ancestors of the superfamily of Hominoidea (Humans and Apes, including Chimpanzees, Gorillas, Orangutans, and Gibbons).
By the end of the Miocene period (6-5 mya), the ancestors of Humans (genus Homo) had split off from the ancestors of Apes. During the Pliocene and Pleistocene periods (5 mya and later), a number of early hominin lines evolved, including Australopithecines, who were ancestors of Homo sapiens.
The Egyptian Fayum
The most productive region for recovering fossils of early anthropoids has been the Fayum region of Egypt, an extensive, low-lying, partly swampy depression located about 80 km south of the Nile Delta. The more arid portions of the Fayum north of Lake Meroe have eroded, badlands exposures of Late Eocene and Oligocene strata including the Jebel Qatrani Formation, containing abundant evidence of the fauna and flora of these periods.
The Fayum was explored in the early 1900s by expeditions of the American Museum of Natural History (Osborn 1908), and several German paleontologists (Markgraf; Schloser 1911). Since the 1960s, investigations of the Jebel Qatrani Formation led by Elwyn Simons
 have greatly increased knowledge of early anthropoids. As Simons describes (1995b), "a
series of expeditions to the Fayum badlands originating first from Yale
(between 1961 and 1968) and then from Duke (from 1977 onward) has
resulted in the discovery of hundreds of mandibles, maxillae, and
postcranials of small primates." The Jebel Qatrani Formation has yielded a
multitude of Eocene and Oligocene primate fossils, placed in 26
species, 19 genera, and 10 families including both prosimians and
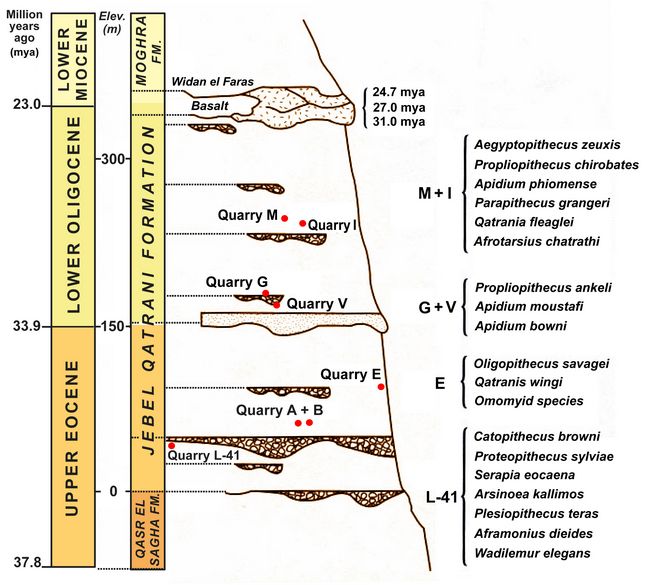
anthropoids (Simons et al. 2007). Quarry I, for example, which has produced many early Oligocene fossils of both Apidium and Parapithecus, was discovered in December 1962 and has been collected from almost continuously since then (Simons 2001). Skulls of Catopithecus, Proteopithecus, Apidum, Parapithecus, and Aegyptopithecus, are described below.
have greatly increased knowledge of early anthropoids. As Simons describes (1995b), "a
series of expeditions to the Fayum badlands originating first from Yale
(between 1961 and 1968) and then from Duke (from 1977 onward) has
resulted in the discovery of hundreds of mandibles, maxillae, and
postcranials of small primates." The Jebel Qatrani Formation has yielded a
multitude of Eocene and Oligocene primate fossils, placed in 26
species, 19 genera, and 10 families including both prosimians and
anthropoids (Simons et al. 2007). Quarry I, for example, which has produced many early Oligocene fossils of both Apidium and Parapithecus, was discovered in December 1962 and has been collected from almost continuously since then (Simons 2001). Skulls of Catopithecus, Proteopithecus, Apidum, Parapithecus, and Aegyptopithecus, are described below.Fig.2: Stratigraphy and quarry locations of fossil anthropoids at the Jebel Qatrani Formation in Egypt (after Simons).
The Jebel Qatrani Formation (fig.2) has been divided into two main faunal zones. The upper zone is dated to older than 31 ± 1 mya from a basalt layer immediately above the formation (Fleagle et al. 1986). The lower zone with a late Eocene green shale unit (Locality 41) has paleomagnetic dating of 36 mya according to Simons et al. (1999), and 34.8-33.9 mya according to Seiffert (2006).
Late Eocene anthropoids
Catopithecus
In 1987 and 1988 fossils of Catopithecus ("below ape"), a previously unknown genus of Anthropoidea, were discovered in Late Eocene strata at site L-41 in the Jebel Qatrani Formation (Simons 1989, 1990, 1995). The Catopithecus fossils resemble those of Oligopithecus from quarry E, and are grouped by Simons with the latter in a subfamily, Oligopithecinae, within the family Propliopithecidae.
The type species, Catopithecus browni (holotype CGM 41885) consists of a right mandible with most teeth and/or alveoli intact, showings a lower dental formula of 2-1-2-3, as in Catarrhini or Old World Monkeys (Simons 1989). Later finds from the same site include a crushed cranium of Catopithecus (fig.3).
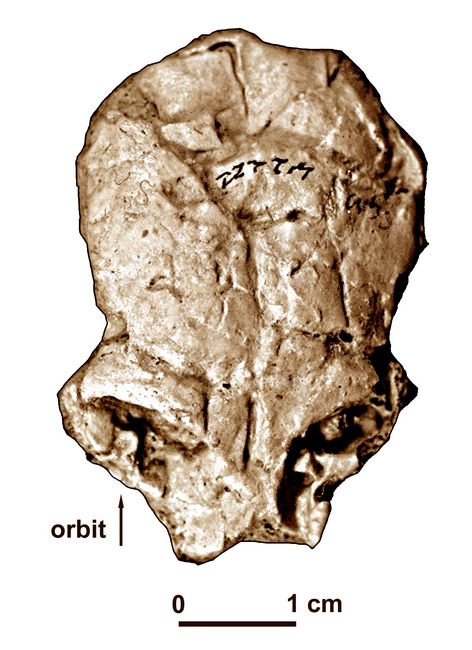 The dentition of C. browni
shows well developed crests which indicate a leaf-eating and/or
insectivorous diet. The relative size of the eye orbits of C. browni
, when compared to those of modern nocturnal and diurnal
anthropoids, indicates that C. browni was
most likely diurnal, comparing most closely to modern prosimians and
callitrichids (marmosets and tamarins, both New World Monkeys; Rasmussen and Simons 1992).
The dentition of C. browni
shows well developed crests which indicate a leaf-eating and/or
insectivorous diet. The relative size of the eye orbits of C. browni
, when compared to those of modern nocturnal and diurnal
anthropoids, indicates that C. browni was
most likely diurnal, comparing most closely to modern prosimians and
callitrichids (marmosets and tamarins, both New World Monkeys; Rasmussen and Simons 1992). Fig.3: Skull of Catopithecus brownii, viewed from top (after Simons et al.)
In terms of evolutionary traits, according to Simons (1990), the Catopithecus skull shows four characteristics of higher primates: a catarrhine dental formula, an ectotympanic at the rim of the auditory bulla, a fused frontal bone, and postorbital closure.
Details of tooth structure (premolars and molars) and a possibly unfused mandibular symphysis resemble those in some Eocene prosimians. According to Simons (1990), the dental anatomy of Catopithecus points to a derivation of Anthropoidea from Eocene adapids rather than omomyids. Evidence for an adapid relationship for Catopithecus are the absence of a tubular ectotympanic, the relatively small incisor alveolae compared to the canine, the possession of lateral incisor alveolae that are larger than those for the central incisors, and similarities of structure in the upper and lower premolars and molars (Simons 1990).
The combination of characters in Catopithecus indicates that it is a very pr
 imitive anthropoidean near the evolutionary
transition from Prosimians. Unlike Prosimiians, however, it has already
acquired frontal fusion, postorbital closure, an ectotympanic in the
lateral wall of the bulla, and a catarrhine dental formula. This set of derived cranial and dental features first occur in
combination in this fossil. Size, shape, orientation of incisors and canines, and other features of the teeth and skull relate Catopithecus both
to later Anthropoidea and to the early and middle Eocene cercamoniine
adapoids (Simons 1995).
imitive anthropoidean near the evolutionary
transition from Prosimians. Unlike Prosimiians, however, it has already
acquired frontal fusion, postorbital closure, an ectotympanic in the
lateral wall of the bulla, and a catarrhine dental formula. This set of derived cranial and dental features first occur in
combination in this fossil. Size, shape, orientation of incisors and canines, and other features of the teeth and skull relate Catopithecus both
to later Anthropoidea and to the early and middle Eocene cercamoniine
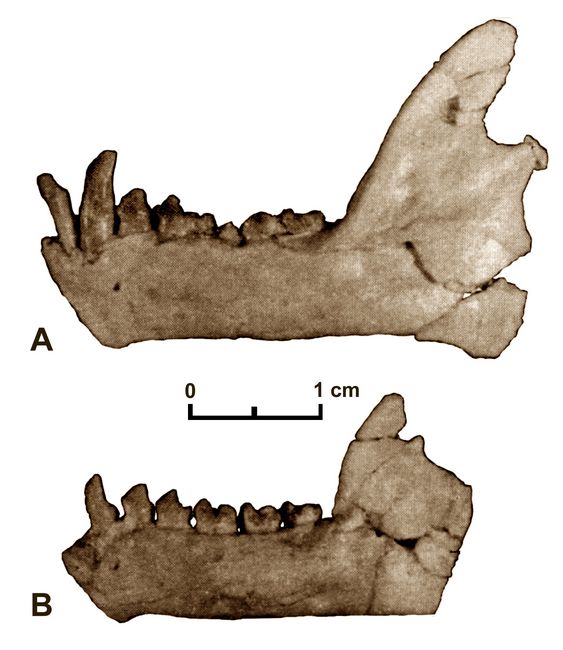
adapoids (Simons 1995).Based on the comparison of mandibles from different individuals (fig.4), sexual dimorphism may be present in Catopithecus (Simons 1995; Simons et al. 1999).
Fig.4: Comparison of (A) left mandible of male Catopithecus DPC 7342 with (B) left mandible of female Catopithecus DPC 11943 (after Simons 1995).
Simons (1995) concludes that the earliest well-documented Anthropoidea occur only in the Fayum, Egypt, and that the retention of some prosimian features by Catopithecus and other Fayum genera indicate they are probably close in time to the initial differentiation of higher primates.
Proteopithecus
Proteopi
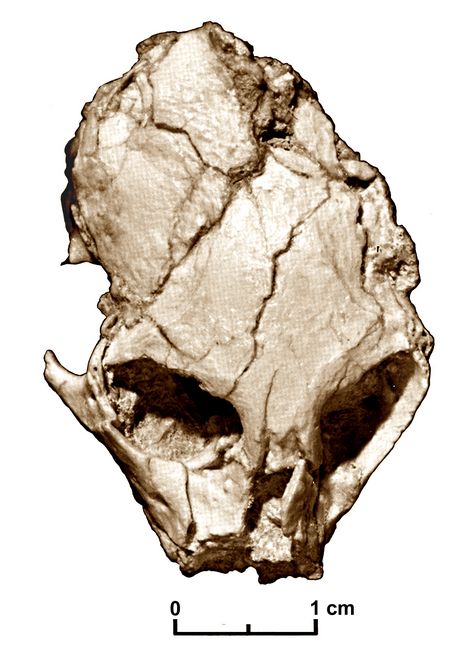 thecus sylviae, found in the Late Eocene L-4l quarry, provides evidence of the Proteopithecidae family, which shows numerous similarities to New World Monkeys. According to Simons (1997), this family
could be part of the basal radiation that produced the New World
platyrrhine primates, or it could be unrelated to any subsequent
lineages. Although no larger than a small callitrichid or a dwarf
lemur, this tiny primate already possessed many of the derived features
of later anthropoids and was a diurnal and probably dimorphic species.
In dental formula and other dental proportions, as well as in known
postcranial features, Proteopithecus more nearly resembles platyrrhines
than does any other Old World higher primate.
thecus sylviae, found in the Late Eocene L-4l quarry, provides evidence of the Proteopithecidae family, which shows numerous similarities to New World Monkeys. According to Simons (1997), this family
could be part of the basal radiation that produced the New World
platyrrhine primates, or it could be unrelated to any subsequent
lineages. Although no larger than a small callitrichid or a dwarf
lemur, this tiny primate already possessed many of the derived features
of later anthropoids and was a diurnal and probably dimorphic species.
In dental formula and other dental proportions, as well as in known
postcranial features, Proteopithecus more nearly resembles platyrrhines
than does any other Old World higher primate. The small size of the Proteopithecus cranium (fig.5) demonstrates that the defining cranial characteristics of Anthropoidea did not arise as a consequence of an increase in size during derivation from earlier prosimians (Simons 1997).
Fig.5: Skull of Proteopithecus sylviae (after Simons 1997, fig.1).
Of all the better known African Paleogene anthropoid primates, Proteopithecus sylviae is the smallest and most generalized (Simons 1997). It exhibits many features that resemble those of platyrrhine (New World) monkeys. General features detectable on the two skulls of Proteopithecus include the relative size and shape of the premaxillae, arrangement and structure of the dentition, position of the lachrymal bone and foramen, degree of postorbital closure, shape of the inferior orbital fissure, closure of the metopic suture, disposition of the temporal lines, arrangement of the palate and nares, and structure of the petrosal area and basicranium.
Although whole or partial crania of the Oligocene genera Aegyptopithecus and Apidium and of the Eocene Catopithecus and Plesiopithecus have been found, most were damaged. Many of the structural details of Proteopithecus are better preserved, including its basicranial anatomy, which can be seen more clearly than in skulls of Plesiopithecus, Aegyptopithecus, or Apidium.
The Proteopithecus cranium (fig.5) is very small, intermediate in size between that of Callithrix jacchus (common marmoset) and Cebuella pygmaea (pygmy marmoset), and approximately the size of crania of Loris tardigradus (slender loris) or Cheirogaleus medius (fat-tailed dwarf lemur). However, orbits of Proteopithecus sylviae are small in comparison with these latter two nocturnal forms (fig.6). In terms of cranial measures, the skull of Aegyptopithecus is approximately 2 to 2 1⁄2 times as large as that of Proteopithecus in linear dimensions. Being contemporaries, Proteopithecus crania can be most closely compared with crania of Catopithecus from L-41.
Taxonomic name Skull len (mm) Width across Orbital ht
orbits (mm) (mm)
Aegyptopithecus zeuxis CGM 40237 104.9 50.6 18.16
Apidium phiomense DPC 9867;AMNH 14556 57.0 est 27.5 est 10.5 est
Proteopithecus sylviae DPC 14095 44.3 25.0 8.5
Proteopithecus sylviae CGM 42214 44.1 est 24.5 est 8.2 est
Catopithecus browni mean of 3 specimens 50.0 28.1 —
Loris tardigradus DPC-O-42 43.6 28.1 14.4
Cheirogaleus medius DPC-O-33 42.8 26.6 11.1
Fig.6: Table comparing cranial sizes in Proteopithecus and other anthropoid primates (after Simons 1997, table 1. Abbreviations: CGM: Cairo Geological Museum; DPV: Duke University Primate Center. AMNH: American Museum of natural History).
Considered overall, the cranial anatomy of Proteopithecus is very distinct from that of strepsirhine prosimians and from that of Tarsius (tarsiers). In dental formula and other dental proportions, as well as in known postcranial features, Proteopithecus more nearly resembles platyrrhines than does any other Old World higher primate (Simons (1997). The hind limb of Proteopithecus is platyrrhine-like, judging from hind limb bones now under study. The broadly rounded posterior jugal of Proteopithecus resembles that of platyrrhines such as Saguinus (tamarin), Callithrix (marmoset), Callimico (Goeldi's marmoset), or Saimiri (squirrel monkey) because of its smoothly rounded surface, resembling the convex side of a spoon, and the temporal lines do not connect across it to form a sharp crest at the top of the zygomatic arch as in most catarrhines (Old World Monkeys).
In Proteopithecus the foramen magnum had occupied a more forward position similar to that seen in Saimiri or Callithrix. In the petrosal region there is a relatively uninflated bulla as in small platyrrhines but unlike the expanded bulla of Tarsius. The postglenoid process is small but distinct. There is a postglenoid foramen that is relatively larger than in Catopithecus and positioned much as in Saguinus or Cebus.
The brain size of Proteopithecus sylviae appears much smaller relative to tooth size when compared with that of a modern small platyrrhine, such as Callithrix. Presumably because of the comparatively small brain, there is more distinct cresting posteriorly, and in this respect the posterior cranium of Proteopithecus resembles that of more robust platyrrhines such as Alouatta (howler monkey), where there is also a distinct occipital crest.
The premaxillary alveolar process, like that of Catopithecus, has a shape and arrangement resembling that of callitrichids (the family including marmosets and tamarins). The frontal process of the maxilla, as in Catopithecus, is rather broad and somewhat vertically arranged. This and other features of the face and front of the skull, which resemble Catopithecus in this region, have been interpreted as indicating a callitrichid-like nasal region with heightened olfactory and scent marking capacities
Proteopithecus stands as representing a separate family from the oligopithecine propliopithecids. This now establishes that there are at least three families of archaic anthropoideans represented in the late Eocene Fayum deposits at L-41: Parapithecidae—Serapia; Propliopithecidae—Catopithecus; and Proteopithecidae— Proteopithecus. If the platyrrhines are to be derived from any known African stock, the only remaining serious candidacy for such a stock is with the family Proteopithecidae (Simons 1997).
Oligocene anthropoids
Apidium
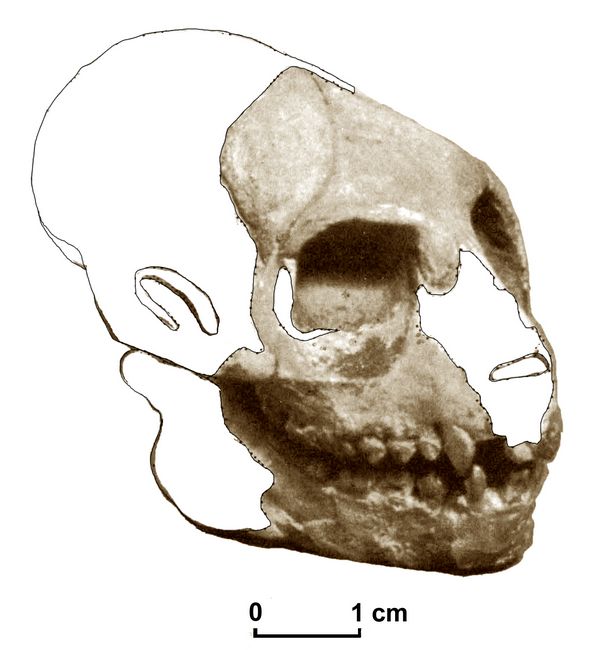
Apidium phiomense represents the most common fossils found in Oligocene levels of the Jebel Qatrani Formation. These mainly occurs in the upper sequence of quarries I and M (fig.3). The type specimen of A. phiomense, a mandible with a fourth premolar and three molars was found by the German paleontologist Richard Markgraf on Feb. 17, 1907 (Simons 1995b).
In 1989 a partial cranium of A. phiomense (DPC) 9867) was located in Quarry I. The specimen is dorso-ventrally crushed and lacks most of the basicranium and part of the facial region (fig.7).
Fig.7: Composite reconstruction of the skull of Apidium phiomense (after Simons 1995)
Parapithecus
Parapithecus fraasi, a primate species described from the Fayum early in this century by Schlosser (1911), has continued to be known only from the type specimen. In 1974, Simons named a second species, Parapithecus grangeri which occurs with the much more commonly found Apidium phiomense at quarries I and M of the Jebel Qatrani Formation (Simons 1995b).
Eventually, a nearly complete skull of Parapithecus grangeri (fig.8) dating from about 33 mya was found in the central part of Fayum Quarry I, a site that lies 200 m above the base of the upper sequence of the Jebel Qatrani Formation (Simons 2001). This skull (DPC 18651) is relatively undistorted, having lost only the upper canines and two pairs of incisors (sockets only) and the left zygomatic arch. It thus represents the most complete higher primate skull yet found in the African Oligocene. Belonging to the superfamily Parapithecoidea, a group regarded by some as the sister group to all other Anthropoidea, this skull reveals important information about the radiation of stem anthropoideans (Simons 2001).
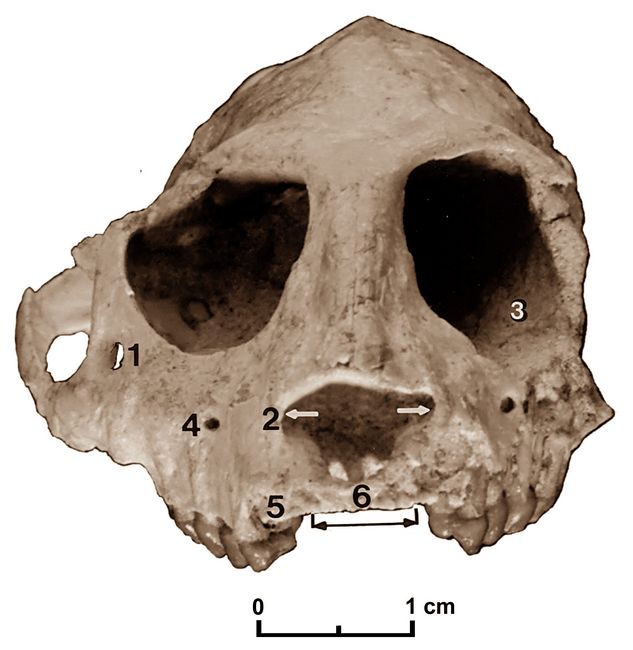
The cranium of Parapithecus is distinctly larger than those of the late Eocene Proteopithecus and Catopithecus, which are only about 70% as long as Parapithecus. In turn, the length of the Parapithecus skull is only about 60% that of the best-preserved Aegyptopithecus skull. The Parapithecus cranium is about the same size as that of the gray gentle lemur, Hapalemur griseus, or of platyrrhines such as the owl monkey, Aotus trivirgatus, or the titi monkey, Callicebus torquatus (Simons 2001).
Fig.8: Skull of Parapithecus grangeri, with several traits labelled: 1.large zygomatico-facial foramen; 2. large nasal aperture; 3.complete postorbital closure; 4. infraorbital foramen; 5. large canine socket; 6. small incisor sockets (after Simons 2001).
Comparatively small orbits and size differences in jaws and teeth show that Parapithecus was both diurnal and sexually dimorphic. This is the only specimen of the species that shows (from sockets) that there were four small upper incisors. Several mandibular examples establish that there were no permanent lower incisors and that the symphysis was fused. Like other early anthropoideans this species possessed a smaller cranial capacity and less-developed orbital frontality than later anthropoideans. There is full postorbital closure and fusion of the metopic suture, and the ectotympanic forms a rim to the auditory aperture (Simons 2001),
The general cranial anatomy of Parapithecus shows very little similarity to that of Tarsiiformes or to strepsirrhines, other than in its comparatively small brain size. Many of the similarities to platyrrhines that can be observed are presumably shared primitive features, such as the absence here of a tubular ectotympanic that is known to be absent also in the early catarrhines Catopithecus and Aegyptopithecus. Unlike tarsiers, omomyids and even Aegyptopithecus, the basisphenoid and basioccipital of DPC 18651 are broad, more as in platyrrhines or even Rooneyia. The mosaic nature of the anatomy of this skull, combining as it does a mixture of primitive, platyrrhine and perhaps even catarrhine characteristics would indicate that cited postcranial resemblance to extant catarrhines are also convergent (Simons 2001).
Aegyptopithecus
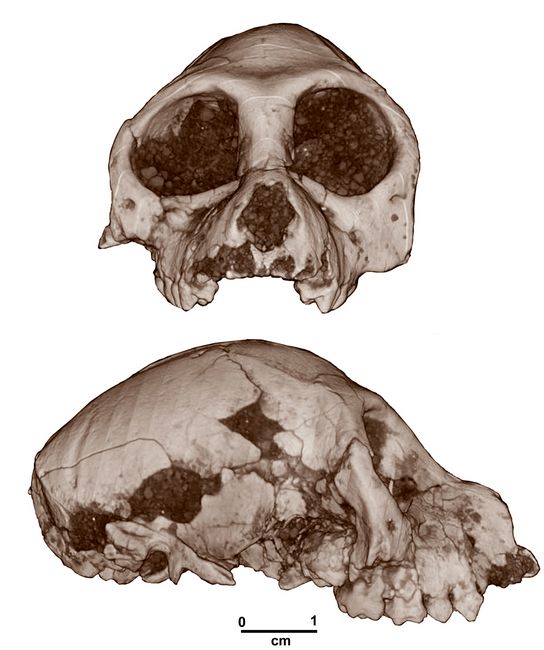
Aegyptopithecus zeuxis is a stem catarrhine species found in the upper levels of the Jebel Qatrani Formation, dating from the Early Oligocene period at 30-29 mya. While a number of Aegyptopithecus and other Propliopithecine skull portions had been previously found, the most complete (and smallest) skull yet known was that of a sub-adult female found in 2004 (Egyptian Geological Museum, Cairo [CGM] 85785). The frontal and right lateral views are shown in fig.9.
 Precise
measurement of this skull shows that Aegyptopithecus had relatively
unexpanded frontal lobes, and a lower brain-to-body mass ratio than
those of living anthropoids. The endocranial volume for CGM 85785, at
14.63 cm3, falls well below previous estimates for A. zeuxis, which had
ranged between 30 cm3 for male skulls and 27 cm3 for female skulls.
Revised measurements of a male skull (CGM 40237) for endocranial volume
by using high-resolution CT scans has yielded a volume of between 20.5
and 21.8 cm3.
Precise
measurement of this skull shows that Aegyptopithecus had relatively
unexpanded frontal lobes, and a lower brain-to-body mass ratio than
those of living anthropoids. The endocranial volume for CGM 85785, at
14.63 cm3, falls well below previous estimates for A. zeuxis, which had
ranged between 30 cm3 for male skulls and 27 cm3 for female skulls.
Revised measurements of a male skull (CGM 40237) for endocranial volume
by using high-resolution CT scans has yielded a volume of between 20.5
and 21.8 cm3.Fig.9: Skull of female Aegyptopithecus zeuxis, CGM 85785 (after Simons et al. 2007)
A significant degree of sexual dimorphism is also indicated in skull size, with the skull of CGM 85785 only 70% the size of a male A. zeuxis skull (CGM 40237), and the latter is even smaller than some other male A. zeuxis crania. Overall, A. zeuxis was highly sexually dimorphic in postcanine tooth size, craniofacial morphology, brain size, and body mass. This is the earliest known example of a peculiar pattern otherwise known only from the fossil record of Miocene catarrhine evolution.
Major differences in the size of male and female Aegyptopithecus led to striking differences in craniofacial morphology. Female crania lacked a well developed rostrum, an elongate sagittal crest, and steep frontal bones. Notably, early and middle Miocene males including Afropithecus turkanensis and Victoriapithecus
macinessi have those features, whereas female specimens such as Proconsul heseloni and Turkanapithecus kalakolensis do not. As Simons et al. (2007) state, further comparisons of craniofacial sexual dimorphism differences with the earlier Aegyptopithecus and other Propliopithecoid taxa should help clarify the phylogenetic relations of these Miocene anthropoids.
..
References:
Fleagle, J.G. 1999. Primate Adaptation and Evolution. Academic Press: San Diego.
Fleagle, J. G., T. M. Bown, J. D. Obardovich, and E. L. Simons 1986. Age of the earliest African anthropoids. Science 234: 1247-1249.
Kay, R.F. J. G. Fleagle, E. L. Simons, Am. J. Phys. Anthropol. 55, 293 (1981);
Osborn, H.F.1908. Bull. Am. Mus. Nat. Hist. 24, 265
Rasmussen, D. T., and E.L. Simons, 1992 . Paleobiology of the oligopithecines, the earliest known anthropoid primates. International Journal of Primatology. 13 (5): 477–508.
Schlosser, M. 1911. Beitr. Paliontol. Geol. Ost.-Ung. 24, 51
Seiffert E.R. 2006. Revised age estimates for the later Paleogene mammal faunas of Egypt and Oman. Proceedings of the National Academy of Sciences. 103 (13): 5000–5005.
Simons, E.L. 1965. Nature 205, 135
Simons, E.L. 1987. J. Hum. Evol. 16, 273
Simons, E.L. 1989.
Simons, E.L. 1990 Discovery of the Oldest Known Anthropoidean Skull from the Paleogene of Egypt. Science
Simons, E.L. 1995a. Skulls and Anterior Teeth of Catopithecus (Primates:Anthropoidea) from the Eocene and Anthropoid Origins. Science
Simons, E.L. 1995b. Crania of Apidium: Primitive Anthropoidean (Primates, Parapithecidae) from the Egyptian Oligocene. American Museum Novitates No. 3124/
Simons, E.L. 1997. Preliminary description of the cranium of Proteopithecus sylviae, an Egyptian late Eocene anthropoidean primate. Proc.Nat.Acad.Sci. no. 94, pp.14970-14975.
Simon, E.L. 2001. The cranium of Parapithcus grangeri , an Egyptian Oligocene anthropoidean primate. Proc.Nat.Acad.Sci. no. 98, pp.7892-7897.
Simons, E.L.,J.M. Plavcan, and J.G. Fleagle 1999. Canine sexual dimorphism in Egyptian Eocene anthropoid primates: Catopithecus and Proteopithecus. Proceedings of the National Academy of Sciences. 96 (5): 2559–2562.
Simons, E.L., E.R.. Seiffert, T.M. Ryan, and Y.Attia 2007. A remarkable female cranium of the early Oligocene anthropoid Aegyptopithecus zeuxis (Catarrhini, Propliopithecidae). Proc.Nat.Acad.Sci. 104, no. 21, pp.8731-8736.
Glossary