Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
22. Cynodonts to Mammaliformes
Cynodonts
Cynodontia (“dog teeth”), grouped as a suborder of the Therapsid order, are considered the direct ancestors of early mammals by the Late Triassic. For a long time, the earliest fossil evidence of mammals did not appear until the Early Jurassic at about 180 mya, in the form of Morganucodons. In the 1990s, however, earlier mammal forms began to appear, such as Adelobasilicus from the Late Triassic in west Texas (Lucas 1990, Lucas and Luo 1993).
In the transition from early reptiles to mammals, various basic changes occurred in the skull, dentition, and post-cranial skeleton. The skull form changed substantially during the process of of accomodating stronger chewing muscles, and of repackaging an expanded brain and special sense organs. Dentition became more specialized, with changes in both teeth and jaws. Limbs and vertebrae changed, related to locomotion with legs placed under the body. Importantly, in conjunction with changes in the jaw anatomy, the mammalian inner ear evolved with three ossicles used as amplifiers, as opposed to a single one in reptiles.
Anatomical traits used to identify mammals, and contrast them with reptiles, are listed in Table 1:
|
Table
1: Major
skeletal differences between early reptiles and mammals. |
||
|
trait |
Early
Reptiles |
Mammals |
|
1 |
No fenestrae in skull |
large
fenestra
exposes all of braincase |
|
2 |
Braincase
attached loosely |
Braincase
attached firmly to skull |
|
3 |
No
secondary
palate |
Complete
bony
secondary palate |
|
4 |
Undifferentiated
dentition |
Incisors,
canines, premolars, molars |
|
5 |
Cheek
teeth
are uncrowned points |
Cheek
teeth
(PM & M) are crowned & cusped |
|
6 |
Teeth
replaced
continuously |
Teeth
replaced
once at most |
|
7 |
Teeth
with
single root
|
Molars
double-rooted |
|
8 |
Jaw
joint made
up of quadrate + articular |
Jaw
joint made
up of dentary + squamosal (*) |
|
9 |
Lower
jaw
composed of several bones |
Lower
jaw
composed of dentary bone only |
|
10 |
Single
middle
ear bone or ossicle (stapes) |
Three
ossicles
in middle ear (stapes, incus, malleus) |
|
11 |
Joined
external nares (nostrils) |
Separate
external nares |
|
12 |
Single
occipital condyle |
Double
occipital condyle |
|
13 |
Long
cervical
(neck) ribs
|
Cervical
ribs
tiny, fused to vertebrae |
|
14 |
Lumbar
(lower
back) region with ribs |
Lumbar
region
rib-free |
|
15 |
No
diaphragm
|
Diaphragm
present |
|
16 |
Limbs
sprawled
out from body |
Limbs under body |
|
17 |
Scapula
simple |
Scapula
with big
spine for muscles |
|
18 |
Pelvic
bones
unfused |
Pelvis
fused |
|
19 |
Two
sacral (hip)
vertebrae |
Three
or more
sacral vertebrae |
|
20 |
Toe
bone numbers
2-3-4-5-4 |
Toe
bone numbers
2-3-3-3-3 |
|
21 |
Body
temperature
variable |
Body
temperature
constant (homeostatic) |
|
(*) note: The
presence of a dentary-squamosal jaw joint has been arbitrarily selected
as the defining trait of a mammal. |
||
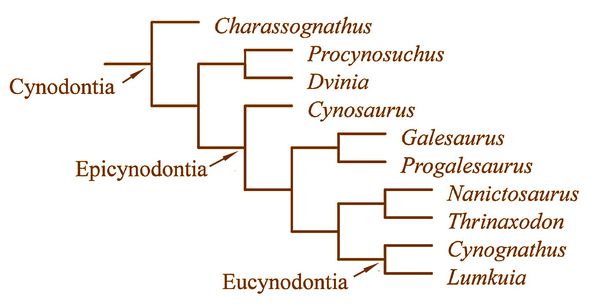
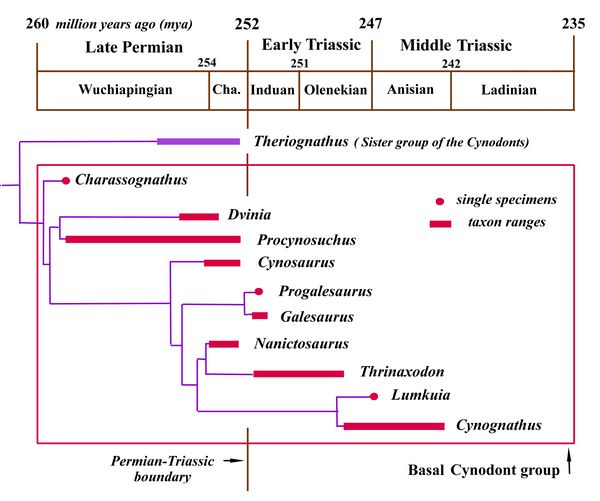
The transition from synapsid reptiles to mammals is well documented, with a long series of fossils linking vertebrate classes and families, permitting the tracing of the origin and radiation of various mammalian orders in some detail (Szalay et al., 1993; Gingerich 1977). While the mammal-like anatomy of cynodonts has been long known, only recent finds have made it appear demonstrable that mammals descended from a single group of cynodonts. This discussion will cover a series of cynodonts leading up through the Eucynodonts ("true cynodonts"), from whom developed the ancestors of mammals by the end of the Triassic. These include Charassognathus, Procynosuchus, Dvinia, Galeosaurus, Thrinaxodon, Cynognathus, and Probainognathus, whose relationships are illustrated in fig.4.
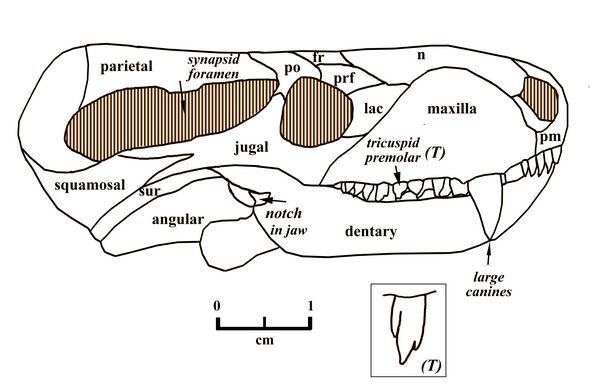
Charassognathus (“notched jaw”), one of the earliest cynodonts, was a very small carnivore, dating from the Late Permian at about 260 mya. The type species, Charassognathus gracilis, was found near Fraserburg, South Africa (Botha et al., 2007). Charassognathus, whose skull was only two inches (5 cm) long, is named for a notch at the back of its lower jaw which was used to attach a chewing muscle called the adductor mandibula externus.
Fig.1: Charassognathus gracilis skull
Charassognathus gracilis also had a the cynodont’s typically large synapsid opening for chewing muscles. Its teeth were diversified into the four cynodont (and mammalian) classes of incisors, canines, premolars, and molars. These included both the very large canines typical of cynodonts, and tricuspid premolars.
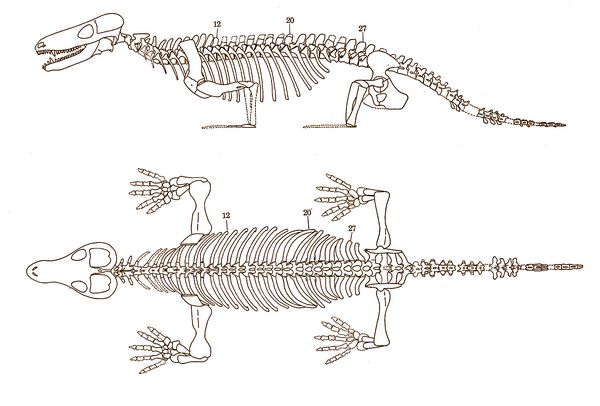
Another early cynodont was Procynosuchus ("before dog crocodile"), who flourished during the Late Permian (260-253 mya). Their fossils, found in the Cistecephalus and Daptocephalus Zones of South Africa and Zambia, and Tartarian Zone IV of Russia, showed they evolved from the same basic therapsid stock as the therocephalians. Procynosuchus was named by South African paleontologist Robert Broom in 1937.
Fig.2: Procynosuchus delaharpe skeleton
Procynosuchus delaharpae, first described by Broom, was an early cynodont with many primitive features, but also some unique skeletal features in the ribs and limbs, some possibly serving as adaptations for semi-aquatic, riverine or estuarine habits. P. delaharpae has been found more re
 cently in the Madumbabisa Mudstone of the Middle Luangwa Valley in
Zambia (Kemp 1979, 1980).
cently in the Madumbabisa Mudstone of the Middle Luangwa Valley in
Zambia (Kemp 1979, 1980). Like some of the therocephalians, the cynodonts possessed a long series of small multicusped cheek teeth and a partial secondary palate. However, the cynodonts also possessed certain unique features as well, such as a closed braincase which probably served generally to strengthen the skull and to protect the brain and middle ear cavity from pressures generated by the contraction of the increased mass of jaw muscles.
Fig.3: Procynosuchus delaharpe skull
They show many advanced, mammal-like characteristics, such as a reduced number of bones in the lower jaw. Even though the jaw hinge remained reptilian, the dentary now made up about 75% of the lower jaw, with the other reptilian lower jaw bones now much reduced, near the jaw hinge. A partially formed, secondary bony palate was composed mainly of palatine bones (a mammalian trait), rather than vomers and maxilla as in reptilian forms.
Procynosuchus also had a complex pattern on the crowns of their cheek teeth. The complex cheek teeth and secondary palate show that the cynodonts were able to chew and breathe at the same time (which most reptiles cannot). The food was broken down in the mouth rather than the stomach. Modifications in the jaw and the flared
 zygomatic arch reflect increased differentiation of jaw musculature.
Lower incisor teeth was reduced to four per side, instead of the
previous six; early mammals, by comparison, had three lower incisors
per side (Kemp 1982). Their vertebral column shows that the
lumbar vertebrae had started to lose ribs, the first sign of functional
division into thoracic and lumbar regions.. A diaphragm may have been present, which would have enhanced
breathing.
zygomatic arch reflect increased differentiation of jaw musculature.
Lower incisor teeth was reduced to four per side, instead of the
previous six; early mammals, by comparison, had three lower incisors
per side (Kemp 1982). Their vertebral column shows that the
lumbar vertebrae had started to lose ribs, the first sign of functional
division into thoracic and lumbar regions.. A diaphragm may have been present, which would have enhanced
breathing.Fig 4: Chart showing the ancestral relationships or phylogeny of cynodonts (after Botha et al. 2007)
Dvinia, another early cynodont from the Late Permian, has been found in Russia in the Upper Tatarian deposits of Arkhangel'sk. Dvinia (named for the Dvinia River in Russia) had cheek teeth with tiny cusps, an initial sign of teeth developing into more than simple perforators.
Dvinia (fig.5) also had an enlarged brain, with various changes occuring in the floor of the braincase. The dentary bone was now the major bone of the lower jaw. The other jaw bones that had been present in early reptiles were reduced to a complex of smaller bones near the jaw hinge. The stapes or columella (the reptilian middle ear ossicle) was recognized as a cartiligenous feature (Tatarinov 1968).
Fig.5: Skull of Dvinia
The postcranial skeleton of Dvinia remains virtually unknown. It is speculated that they had a higher metabolic rate that may have approached homeothermy (Battail and Surkov 2000; Hopson 1991; Hopson and Barghusen 1986; Kemp 1982; Sidor and Hopson 1998; Tatarinov 1968).
 Galesaurus planiceps (fig.6) is a relatively advanced Cynodont from the Early
Triassic of South Africa, now placed in the clade calleed
Epicynodontia. When first discovered in South Africa and classified by Richard Owen in 1859, Galesaurus was incorrectly identified
as a dinosaur, of which numerous examples were being found in that
region. Ten years later, in 1869, when the fossil genus
Galesaurus was described in the first issue of Nature, it was still thought by T. H. Huxley to be a dinosaur.
Eventually, it was reclassified as a cynodont. As Owen and Huxley
were two of the most knowledgable scientists of
their day, the misidentifaction illustrates problems of classification then caused by a lack
of comparative data.
Galesaurus planiceps (fig.6) is a relatively advanced Cynodont from the Early
Triassic of South Africa, now placed in the clade calleed
Epicynodontia. When first discovered in South Africa and classified by Richard Owen in 1859, Galesaurus was incorrectly identified
as a dinosaur, of which numerous examples were being found in that
region. Ten years later, in 1869, when the fossil genus
Galesaurus was described in the first issue of Nature, it was still thought by T. H. Huxley to be a dinosaur.
Eventually, it was reclassified as a cynodont. As Owen and Huxley
were two of the most knowledgable scientists of
their day, the misidentifaction illustrates problems of classification then caused by a lack
of comparative data. Fig.6: Galesaurus planiceps (after xxx)
Galesaurus had a broad snout, and an enlarged zygomatic arch. Its frontal bones had anterior "spine" projections inserting between nasals, a trait which persisted in adults. It had a complete secondary palate. Its skeleton had distinct lumbar and thoracic regions, with ribs restricted to the thoracic region. A diaphragm was probably completely established in the species. Its limbs were relatively upright (Hopson & Barghusen 1986; Kemp 198; Sidor & Hopson 1998)
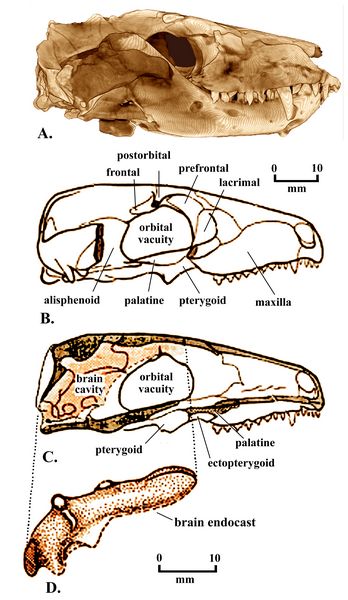
Thrinaxodon liorhinus (fig.7), dating from the early Triassic (252-247 mya), was a badger-like carnivore about 50 cm long, with short limbs and a short tail. Most examples have been discovered in South Africa, and are in museums in Cape Town, Bloemfontein, and Johannesburg. The discovery by Edwin Colbert of Thrinaxodon in Antarctica during the 1960's provided a definite indication that Antarctica and Africa were connected as part of Pangaea during the Early Triassic.
T. liorhinus was a relatively advanced "galesaurid" cynodont, with development of several of the cynodont features. Its temporal fenestra was large, with large jaw muscle attachments. Its snout was about as broad as tall, with a narrow braincase and a sharp parietal crest and a robust zygomatic arch. It had a fully ossified secondary palate. It had a near-mammalian functional division of teeth, with incisors (four uppers and three lowers), canines, and 7-9 cheek teeth with cusps for chewing.
Fig.7: Thrinaxodon liorhinus, skull and brain endocast..
The brain (fig.7) was long and narrow, resting within a very narrow skull. Like other cynodonts the brain filled the endocranial cavity (Rowe 1996). In Thrinaxodon, the braincase no longer needed to support muscle. This allowed expansion of the braincase, which in turn increased the surface area available for muscle attachment (Palaios 2014).
The more reptilian or primitive features of Thrinaxodon include the lower jaw attachment, with only primary evidence of a dentary-squamosal articulation. The Thrinaxodon dentary, however, was large, with the little quadrate and articular bones now loosely attached. The stapes now touched the inner side of the quadrate. The first si
 gn of the
mammalian jaw hinge appeared, in the form of a ligamentous connection
between the lower jaw and the squamosal bone of the skull. The
occipital condyle now consisted of two slightly separated surfaces,
though not as far separated as the mammalian double condyles (Hunt
1994-7). It retained a pineal or "third eye" opening in the skull.
gn of the
mammalian jaw hinge appeared, in the form of a ligamentous connection
between the lower jaw and the squamosal bone of the skull. The
occipital condyle now consisted of two slightly separated surfaces,
though not as far separated as the mammalian double condyles (Hunt
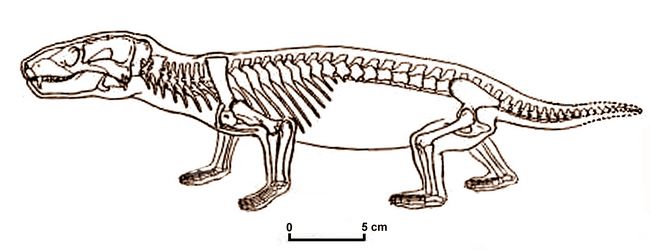
1994-7). It retained a pineal or "third eye" opening in the skull. Fig.8: Skeleton of Thrinaxodon (after Jenkins 1984).
Its teeth had a "reptilian" alternate replacement pattern. Its cheek teeth had multiple cusps but were all alike (i.e., no distinction between molars and premolars). Thrinaxodon’s vertebral connections were more mammalian, and all four legs stood fully upright, not sprawling.
Fig 9: Chart showing chronology of cynodonts (after Botha et al. 2007).
Eucynodonts
Eucynodontia ("true dog teeth") is an infraorder of Therapsids that includes both mammals and most non-mammalian cynodonts. The oldest eucynodonts are known from the beginning of the Early Triassic, and possibly the end of the Late Permian. Eucynodontia includes two major subgroups, Cynognathia and Probainognathia.
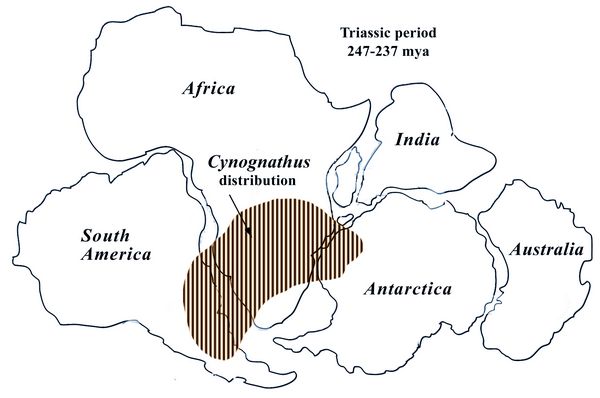
 Cynognathus (“dog jaw”) was a meter-long carnivore closely related to
mammals that lived during the Early and Middle Triassic (247-237 mya).
It is known from a single species, Cynognathus crateronotus, which had
a very wide distribution in Gondwanaland, the southern part of Pangaea
(fig.11). Fossils have been found in the Karoo formation of South
Africa; the Fremouw Formation, in South Africa/Lesotho; in the Puesto
Viejo Formation in Argentina; in Antarctica; and in China.
Cynognathus (“dog jaw”) was a meter-long carnivore closely related to
mammals that lived during the Early and Middle Triassic (247-237 mya).
It is known from a single species, Cynognathus crateronotus, which had
a very wide distribution in Gondwanaland, the southern part of Pangaea
(fig.11). Fossils have been found in the Karoo formation of South
Africa; the Fremouw Formation, in South Africa/Lesotho; in the Puesto
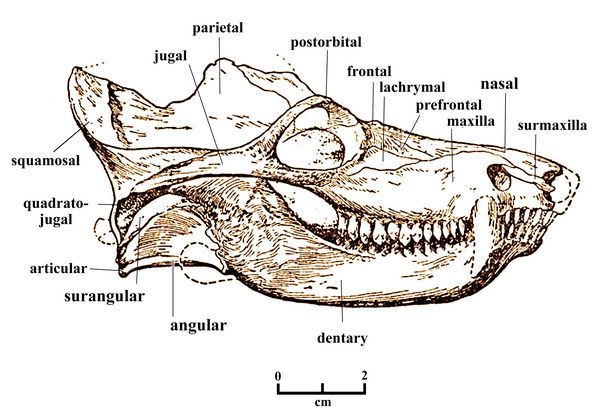
Viejo Formation in Argentina; in Antarctica; and in China.Fig.10: Skull of Cynognathus crateronotus
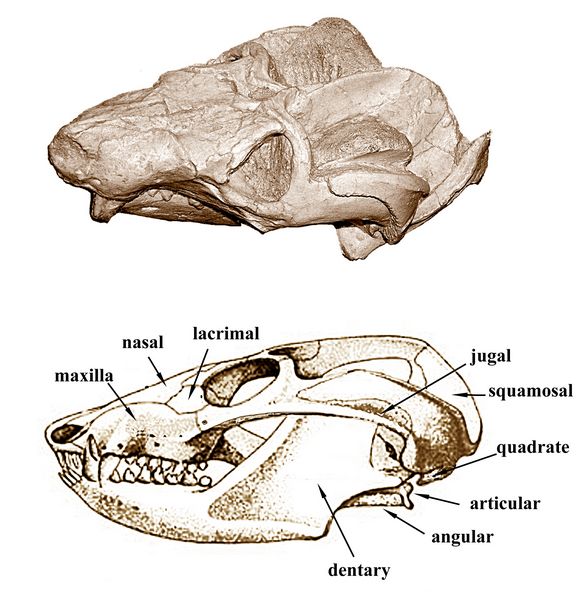
Cynognathus (fig.10)was a heavily built animal, with a large head about 30 cm (1.0 ft) in length, wide jaws, and sharp teeth. The dentary (lower jaw) was equipped with differentiated teeth, and the maxilla or upper jaw included large canines, both traits typical of cynodont carnivores. The temporal fenestra was large, indicating a strong chewing muscle. The cheek teeth had cusps which met in true occlusion for more efficiently slicing up food and the rate of tooth replacement was reduced, with mammalian-style tooth roots (although still single roots). Pits and canals on the premaxilla or nasal bone represent concentrations of nerves and blood vessels. In mammals, such structures allow hairs or whiskers to be used as sensory organs.
 The
dentary now comprised about 90% of the muscle-bearing part of the lower
jaw. It contained a transitional jaw joint, combining mammalian and
reptilian features. A new bony jaw joint existed between the squamosal
(skull) and the surangular bone (lower jaw), while the other jaw joint
bones were reduced to a compound rod lying in a trough in the dentary,
close to the middle ear.
The
dentary now comprised about 90% of the muscle-bearing part of the lower
jaw. It contained a transitional jaw joint, combining mammalian and
reptilian features. A new bony jaw joint existed between the squamosal
(skull) and the surangular bone (lower jaw), while the other jaw joint
bones were reduced to a compound rod lying in a trough in the dentary,
close to the middle ear. Fig.11: Distribution map of Cygnognathus crateronotus
The presence of a secondary palate in the mouth indicates that Cynognathus would have been able to breathe and swallow simultaneously. Its hind limbs were placed directly beneath the body, but the forelimbs sprawled outwards in a reptilian fashion. Its ribcage formation was more mammalian, with ribs now lacking in the stomach region, suggesting an efficient diaphragm, an important muscle for mammalian breathing.
The descendents of Cynognathus gradually evolved into small carnivorous and insectivorous forms, perhaps nocturnal, depending on their senses of hearing and smell. Due to their enlarged olfactory and auditory bulbs, Cynodont and early mammalian brains gradually grew larger than sauropsid (reptilian) brains.
Regarding the development of the mammalian middle ear, the eardrum had developed in the lower jaw, behind the jaw hinge, supported by a wide prong (reflected lamina) of the angular bone. Airborne sound was transmitted through the eardrum to two small lower jaw bones, the articular and the quadrate, which contacted the stapes in the skull, which contacted the cochlea.
Cynodonts, who essentially had two jaw joints, developed relatively loose quadrates and articulars that could vibrate freely for sound transmittal while still functioning as a jaw joint, strengthened by the mammalian jaw joint right next to it. All early mammals from the Lower Jurassic retained this low-frequency ear and a double jaw joint. By the middle Jurassic, mammals lost the reptilian jaw joint (though it still occurs briefly in embryos) and the articular and quadrate bones became miniaturized and migrated into the nearby middle ear as the incus and malleus, serving to amplify high-frequency sounds.
Probainognathus (fig.12), an advanced cynodont from the Middle Triassic (239-235 mya), was first found in Argentina, and later in South Africa, with the related genus Lumkuia. More recently Probainognathian fossils from the Middle Triassic have also been found in Madagascar. There they were part of an assemblage dated at 230-225 mya, that included early dinosaur fossils as well as several other cynodonts (Flynn et al 1999.; Carroll 1988).
The Probainognathians, like the Cygnognathians, were mostly carnivorous, with jaws considerably shorter than those of Cygnognathus, but with similar tooth forms including large canines. Probainognathians are split into four families: Probainognathidae, Chiniquodontidae, Tritheledontidae, and Mammaliaformes (the latter group comprising ancestors of mammals). Non-mammalian probainognathians, one of the longest-lived therapsid groupings, lived from the Triassic to the Jurassic. Probainognathians spread across the Gondwana of Triassic times, although not as widely as
 Cygnognathus.
Cygnognathus. Probainognathus had a relatively large brain showing various skull changes and the fusion of some skull plates. One of these involved the closing of the pineal, "third eye" formamen. The slender cheekbone was relatively low on the side of the eye socket, although the postorbital bar (a reptilian trait) was still present. In terms of digits, its had the same number of toe bones as mammals.
The evolution of chewing muscles in late synapsids is well illustrated by Probainognathus, showing increasing muscular control related to grinding functions of specialized molars. This control involved a split masseter or cheek muscle which had more distributed attachments (Palaios). Undoubtely this was linked to additional cusps on the molars or cheek teeth of Probainognathus.
Fig.12: Probainognathus skull
End of the therapsid carnivores
While some cynodont groups lasted well into the Jurassic period, many therapsid carnivores became extinct during the multi-stage Permian–Triassic extinction event. This effectively ended the dominance of the carnivores among the therapsids. In the early Triassic, all the medium-to-large land carnivore niches were taken over by archosaurs (diapsid reptiles) which, over about 35 million years, came to include the crocodylomorphs, the pterosaurs, and the dinosaurs. By the Jurassic, the dinosaurs had come to dominate the large terrestrial herbivore niches as well.
By Early Triassic times, cynodonts had diverged into large predaceous carnivores such as Cynognathus and moderate large omnivorous and herbivorous types such as Trirachodon and Diademodon. The Middle Triassic saw a major radiation of herbivorous forms included in the family Traversodontidae. From this family evolved the highly specialized and extremely mammal-like Tritylodontidae of the Late Triassic to Middle Jurassic, the "rodents" of the early Mesozoic and culmination of the herbivorous cynodont radiation.
By the late Triassic, the Therapsid carnivores had become small, about the size of a weasel or rabbit. The first mammals, also small in size, appeared in the Late Triassic epoch at about 220 mya, some 50 million years after the first therapsids. The majority of the mammal species that existed in the Mesozoic Era were nocturnal insectivores, the direct ancestors of multituberculates and primates.
.
.
References:
Barghusen (1968)
Battail & Surkov 2000;
Benton, M. J. 2000 , Vertebrate Paleontology, 2nd Ed. Blackwell Science Ltd (2004) 3rd edition
Berman, D.S. 1977. "A New Species of Dimetrodon (Reptilia, Pelycosauria) from a Non-Deltaic Facies in the Lower Permian of North-Central New Mexico". Journal of Paleontology 51(1): 108—115;
Carroll, R. L. 1988, Vertebrate Paleontology and Evolution, WH Freeman & Co
Crompton & Parker (1978);
Flynn et al. 1999, Science
Hopson 1991
Hopson, J.A. and Barghusen, H.R. (1986), An analysis of therapsid relationships in N Hotton, III, PD MacLean, JJ Roth and EC Roth, The Ecology and Biology of Mammal-like Reptiles, Smithsonian Institution Press, pp. 83–106
Hunt, Kathleen 1994-1997 Transition from amphibians to amniotes; transitional Vertebrate Fossils
Huttenlocker, A. K.; Mazierski, D. and Reisz, R. R. 2011. "Comparative osteohistology of hyperelongate neural spines in the Edaphosauridae (Amniota: Synapsida)". Palaeontology 54: 573–590.)
Ivakhnenko, M.F. 1999, Biarmosuches from the Ocher Faunal Assemblage of Eastern Europe, Paleontological Journal vol 33 no.3 pp. 289–296. abstract
Kemp, T.S. 1980. "The primitive cynodont Procynosuchus: structure, function and evolution of the postcranial skeleton". Philosophical Transactions of the Royal Society B 288 (1027): 217–258.
Kemp (1982);
Lucas, S.G.; Spielman, J. A.; Rinehart, L.F.; Martens, T. (2009). "Dimetrodon (Amniota: Synapsida: Sphenacodontidae) from the Lower Permian Abo Formation, Socorro County, New Mexico". New Mexico Geological Society Guidebook, 60th Field Conference, Geology of the Chupadera Mesa Region: 281–284.)
Lucas, S. G. (2011). Traces of a Permian Seacoast. Prehistoric Trackways National Monument. pp. 1—48.)
Marsh, O.C. (1878). "Notice of new fossil reptiles". American Journal of Science 3(15): 409-41)
Palaios, 2013, Mammals (pdf. file including various articles from their web site at www.palaios.org)
Romer, A.S.; and Price, L.I. (1940). "Review of the Pelycosauria". Geological Society of America Special Paper 28: 1–538.
Rowe 1996
Sidor & Hopson (1998);
Tatarinov, L.P. 1968. Morphology and Systematics of the Northern Dvinia Cynodonts (Reptilia, Synapsida; Upper Permian). Yale University Postilla 126.
Glossary