Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
11. Lobe finned fish to Tetrapodomorphs
The class of Osteichthyes or bony fish is divided into two main subclasses, the ray finned fish or Actinopterygians, and the lobe-finned fish or Sarcopterygians. The ray finned fish include most well-known marine and freshwater fish, ranging from ancient types such as gar (Lepisosteus), with ganoid or enamel-covered scales, to more recent groupings with scales such as today’s salmon and trout. The Sarcopterygians or lobe-finned fish, meanwhile, although with very few species left today, are the ancestors of all land vertebrates (tetrapods), who first appear as tetrapodomorph fishes.
Sarcoptygerians.
Early lobe-finned fishes have fleshy, lobed, paired fins, which are joined to the body by a single bone. Their fins differ from those of all other fish in that each is borne on a fleshy, lobelike, scaly stalk extending from the body. Pectoral and pelvic fins have articulations resembling those of tetrapod limbs. These fins evolved into legs of the first tetrapod land vertebrates, amphibians. The Sarcopterygians fins have articulating bones, contrasting with the thin, spiny fins of Actinopterygians or ray-finned fish. The lobe fins rotate on shoulder or hip sockets, like the limbs of tetrapods and all later land vertebrates. This includes the individual rays in the lobe fin, which can be moved individually, like fingers and toes. Most of the Sarcopterygians’ fin bones directly correlate with limb bones of land animals.
Sarcoptygerians also possess two dorsal fins with separate bases, as opposed to the single dorsal fin of actinopterygians (ray-finned fish). The braincase of sarcoptergygians primitively has a hinge line, but this is lost in tetrapods and lungfish. Many early lobe-finned fishes have a symmetrical tail. All lobe-finned fishes possess teeth covered with true enamel.
The Sarcopterygii first appeared by the Late Silurian period. The earliest well-known forms are from the Early Devonian and include forms like Youngolepis, and Powichthys, whose affinities are still undetermined. They are variously considered as primitive proto-lungfish, or ancestral or "stem-group" sarcopterygians from which the other forms evolved. The largest known lobe-finned fish was Rhizodus hibberti, up to 7 m long, from the Carboniferous period of Scotland.
In the Early Devonian period (416 - 397 mya), the sarcopterygians split into two main lineages, the coelacanths and the rhipidistians.
Coelacanths.
The coelacanths, once an important group, had their greatest expansion during late Devonian and Carboniferous, from 385 to 299 mya. At the end of the Devonian period, a major extinction event occurred with elminated most marine forms of sarcoptyerygians, with only the coelacanths surviving. Freshwater types, however, including lungfish and thizodonts, remained more plentiful. Rhizodonts, large predators who grew up to several meters long, lived from the Late Devonian through Pennsylvanian periods, then became extinct.
Coelocanths are now limited to the single large deepwater genus Latimeria.
Rhipidistia.
The Rhipidistians, whose ancestors probably lived in the oceans near the river mouths or estuaries, left the salt water, marine world and migrated into freshwater habitats. They, in turn, split into three orders: a) the lungfish (Dipnoi), of which a few species still survive; b) the Porolepiformes, closely related to lungfish, and possibly themselves ancestral to some amphibians; and c) the tetrapodomorphs, including the ancestors of tetrapods, the first land vertebrates. The Dipnoi (lungfish) and Porolepiformes are together gouped as Dipnomorpha (Jarvik 1997).
Dipnomorpha.
The order Dipnoi (lungfish)
Lungfish (Dipnoi) are bony fish with primitive lungs as well as gills. As Sacroptygerians, lungfish have also lobed fins with a well-developed internal skeleton. Lungs and lobed fins represent traits that led to tetrapods and amphibians. The lungfish evolved the first proto-lungs and proto-limbs; developing the ability to live outside a water environment in the middle Devonian (397 - 385 Ma). The lungfish's greatest diversity was in the Triassic period; today there are fewer than a dozen genera left.
All Osteichthyians (bony fish) possess gills. For the majority this is their sole or main means of respiration. Osteichthyian also typically have swim bladders, which helps the body float. In the case of lungfishes, vascularized swim bladders have developed into primitive lungs. Lungfish can breathe air and survive on land. While other species of bony fish can breathe air via modified, vascularized gas bladders, these are usually simple sacs, devoid of complex internal structure. In contrast, the lungs of lungfish are subdivided into numerous smaller air sacs, maximizing the surface area available for gas exchange. All lungfish have two lungs, with the exception of the Australian lungfish, which only has one. The lung(s) connect to the pharynx. The lungs of lungfish are homologous to the lungs of tetrapods. In each case, the lungs extend from the ventral surface of the esophagus and stomach.
Lungfish are omnivorous, feeding mainly on bottom dwelling invertebrates (crustaceans, worms, molluscs) and plant matter, but also on fish, insects, and amphibians. They remained important as inhabitants of rivers and lakes through the Mesozoic era. The few surviving species of lungfish today in Australia, South America and Africa remain little changed from the Mesozoic. Among these is the genus Neoceratodus from Queensland, Australia. Fossils of nearly identical lungfish have been found in northern New South Wales, showing that today’s Queensland lungfish are virtually unchanged from their fossil ancestors. Neoceratodus represents the most primitive surviving member of ancient air-breathing lungfish, and one of the oldest known living vertebrate genera.
African and South American lungfish are capable of surviving seasonal drying out of their habitats by burrowing into mud and estivating throughout the dry season. This burrowing ability is related to their abilities to dig with their limb-like lobed fins. Burrowing has also been identified for at least one group of fossil lungfish, the Gnathorhizidae, who lived from the Pennsylvanian to Mid Triassic periods. Gnathorhizid fossils are found primarily in levels represting ancient wetlands, often in association with burrow-like structures, suggesting they may have estivated (hibernated) during the dry season. Unlike most fossil lungfish, but like modern South American and African lungfish, gnathorhizids have bladelike toothplates, suggestng they were predators,
The order Porolepiformes
Porolepiformes (“scale pores”) are an extinct order of lobe-finned fish from the Devonian period (416 to 359 mya). The group contains two families: Holoptychiidae and Porolepididae. Their name comes from distinctive rows of pores visible on the scales of some genera (e.g. Porolepis), which are useful as stratigraphic markers.
The order Porolepiformes was established by the Swedish paleontologist Erik Jarvik (1980), who based on the shapes of their snouts, proposed they gave rise to the salamanders and caecilians independently of the other tetrapods, a view not widely shared at present (Schultze and Trueb 1991). Recent phylogenetic reconstruction places porolepiformes close to lungfishes (Janvier 1996). The Porolepiformes and lungfish are grouped together as Dipnomorpha, as a sister group to Tetrapodomorpha. Together, these form the Rhipidistia, the sister group to the Coelacanths.
The Porolepiformes were large predators, with long fins, short broad heads, and small eyes. They represent a somewhat primitive lineage of rhipidsitians. The lower jaw contains several rows of large teeth on the tooth whorl. They are first known as marine fish during the Early Devonian.
Powichthys, from the Early Devonian in North America, was relatively small (less than 30 cm or 1 ft). Its name (“PoW-fish) is derived from an abbeviation for Prince of Wales Island, located in arctic Canada (Chang & Smith 1992; Long 1995). In the Middle Devonian, Porolepiformes successfully colonized brackish and freshwater locales, and flourished through the Late Devonian, especially the cosmopolitan genus Holoptychius. In 2011, a new North American genus of porolepiformes was discovered from the Late Devonian, a large predatory fish named Laccognathus.
Tetropodomorphs.
The Tetrapodomorpha are a now continually redefined group of vertebrates, containing several groups of related lobe-finned fishes, including the orders of Rhizodonts, Tristichopterids, and Elpisosteglia. Essentially, tetrapodomorphs are the closest sarcopterygian relatives of tetrapods, defined by cladistic trait-counting as those that are more closely related to living tetrapods than to living lungfish.
Tetrapodomorph fossils are known from the early Devonian onwards. The first tetrapodomorphs, which included the gigantic rhizodonts, had the same general anatomy as the lungfish, who were their closest kin, but they appear not to have left their water habitat until the late Devonian epoch (385 - 359 Ma), with the appearance of tetrapods (four-legged vertebrates). Tetrapods are the only tetrapodomorphs which survived after the Devonian. Non-tetrapod sarcopterygians continued until towards the end of Paleozoic era, suffering heavy losses during the Permian-Triassic extinction event (251 Ma).
Among the physical traits defining tetrapodomorphs are modifications to the fins, notably a humerus with convex head articulating with the glenoid fossa (the socket of the shoulder joint). More advanced forms like the genus Tiktaalik have sometimes been referred to as "fishapods," appearing in their skull anatomy and limb morphology to be half-fish and half-tetrapods. A more all-embracing definitioon of Tetrapodomorpha includes the crown group tetrapods, the last common ancestor of living tetrapods and all of its descendants.
Rhizodonta.
Rhizodonts (Order Rhizodontida) were a group of predatory tetrapodomorphs living in tropical rivers and freshwater lakes around the world, from the late Devonian (ca. 377 mya) to the Late Pennsylvanian (310 mya). Rhizodonts were the dominant predators of their age, with a set of large tusks and fangs as well as smaller teeth. They reached sizes up to 7 meters long, as in the case of Rhizodus hibberti from Europe and North America, the largest freshwater fish ever known. Their prey may have included sharks, lungfish, other lobe-finned fishes, and early tetrapods, who had had to lay their eggs in water.
Trisichopterida.
Tristichopterids were a diverse and successful group of tetrapodomorph fishes living in the Middle and Late Devonian. Their sizes ranged from about 30 cm, in the case of Tristichopterus, up to several meters in length in genera such as Hyneria and Eusthenodon. All tristichopterids went extinct by the end of Late Devonian.
Some tristichopterids share some of the features of the elpistostegalians, a paraphyletic assemblage of fishes close to the origin of tetrapods. This mainly concerns the shape of the skull and a reduction in size of the posterior fins.
Fossil genera of Tristichopterids include the following: Cabonnichthys; Canningius; Edenopteron; Eusthenodon; Eusthenopteron; Hyneria; Jarvikina; Langlieria; Mandageria; Notorhizodon; Platycephalichthys; Tristichopterus; and Heddleichthys.
Tristichopterus alatus, the type genus, was first named in 1861 (Egerton).
Eusthenopteron
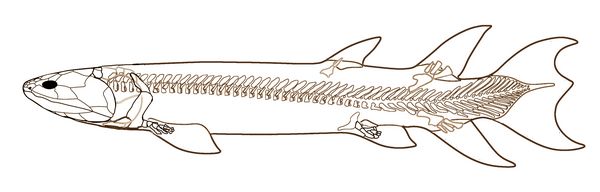
Probably the best known genus of Tristichopterids is Eusthenopteron (fig.5), with several species identified from the late Devonian at about 385 mya. Eusthenopteron was a strictly aquatic animal, although it has at times (erroneously) been portrayed as a candidate for the first fish to walk up on land. Eusthenopteron was first described by J. F. Whiteaves in 1881 from numerous fossils found at Miguasha, Quebec. One Eusthenepteron fossil from Miguasha was studied and described in great detail during the 1940s- 1990s by paleoichthyologist Erik Jarvik.
Eusthenopteron, which grew up to 1.8 m (6.5 ft) in length, shared many unique anatomical features including skull and limb bones with the earliest known tetrapods, such as Acanthostega. and Ichthyostega. It also had la
 byrinthodont teeth, found in all of the earliest known
tetrapods and persisting in amphibians through the Carboniferous
(Laurin et al 2007)
byrinthodont teeth, found in all of the earliest known
tetrapods and persisting in amphibians through the Carboniferous
(Laurin et al 2007)Fig.5: Skeleton of Eusthenopteron fordii (after Jarvik 1980).
While a humerus and femur are present in the fore-fins of all sarcoptergyians, Eusthenepteron had a much more complete set of tetrapod-like fin bones, include a distinct humerus, ulna, and radius in the fore-fin, and a femur, tibia, and fibula fin the pelvic or rear fin(fig.5). Also as in tetrapod long bones, the limb bones of Eusthepteron had epiphyseal growth plates enabling longitudinal growth through ossification.
Elpisosteglia
A third family of Tetrapodomorphs is called Elpisosteglia. This family includes two of the most important tetrapodomorphs, Panderichthus and Tiktaalik.
.
Panderichthys
Panderichthys was a fish about 1 meter long, dating from the Late Devonian period (380 mya). It was found in Latvia and is named after the 19th century German-Baltic palaeontologist and biologist Christian Heinrich Pander. Panderichthys had a large, elongated, tetrapod-like head, and exhibits various transitional features between lobe-finned fishes and early tetrapods such as Acanthostega. In cladistic terms, Panderichthys is considered the most "crownward" stem fish-tetrapod with paired fins.
Of the many anatomical changes required for the transformation from lobe-finned fish to land dwelling tetrapods, probably the most important was strengthening of the limbs and their supporting structures, the pectoral and pelvic girdles (comprising the scapular and innominate bones). CT scans of well-preserved fossils of Panderichthys fossils show four distinct distal radial bones or digits at the end of the fin skeletal structure. While these digits are short and have no joints, they show an intermediate form between the lobe fins of Sarcoptyerigian fish, and the limbs of tetrapods.
In Panderichthys the fin to limb transition began in the pectoral or shoulder appendages, and only later occurred in the pelvic appendage. Its pectoral girdle shows derived characteristics, while its pelvic girdle retains ancestral ones. The shift to legs as weight-bearing limbs and related strengthening of the pelvic area occurred in later tetrapods. While Panderichthys does not show this shift to leg and pelvic strengthening, it seems to have been capable of some kind of shallow water or terrestrial movement, and had the ability to prop itself up.
Tiktaalik
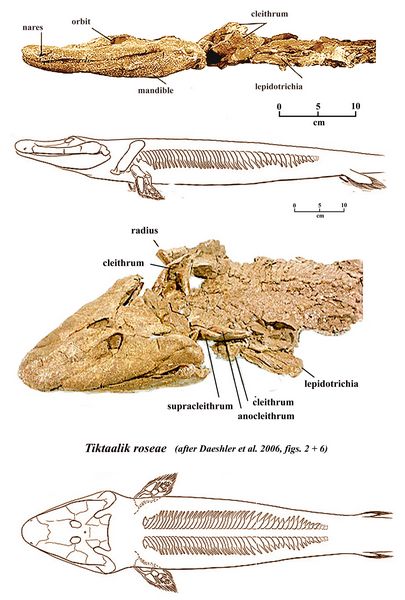
Tiktaalik roseae was a lobe-finned fish and tetrapodomorph dating from the Late Devonian period at 374-356 mya. Tiktaalik, discovered in southern Ellesmere Island in Nunavut Territory, in the Arctic region of Canada, belongs to the order Elpistostegalia, considered more closely related to tetrapods than osteolepiform fishes (Camp and Allison 1961).
Tiktaalik is an Inuktitut word meaning burbot, a freshwater fish related to the true cod. The holotype fossil of the type species T. roseae (NUFV 108) consists of the skull and pectoral (shoulder) region, and the remainder of the skeleton has also been recovered (fig.6). The pectoral fin of the skeleton is composed of bones homologous to the humerus, ulna, and radius of the tetrapod, followed by a host of smaller bones anchoring the fin rays. The pelvic fin skeleton similarly has femur, tibia, and fibula. The body length is 1-3 m. Its neck and ribs were similar to those of tetrapods, with the ribs being used to support its body and aid in its breathing via lungs (Daeschler et al. 2006).
Fig.6: Skeleton of Tiktaalik roseae (after Daeschler et al. 2006, figs. 2+6)).
Tiktaalik was a shallow-water fish which had evolved a shift in propulsion apparatus, from tail fins to pectoral and pelvic fins, and a reliance on lungs rather than gills. Both were adaptations to inland freshwater zones. Tiktaalik had rows of sharp teeth, and well developed jaws suitable for catching prey, both indicating a predator fish. Tiktaalik is considered one of the best candidates for the earliest known tetrapod-like fish who was the ancestors of all land animals.
Tetrapodomorph fish like Eusthenopteron, Panderichthys, and Tiktaalik were the direct ancestors of the first tetrapods, the air-breathing, terrestrial animals from which the land vertebrates, including humans and other mammals, are descended.
.
References:
Camp, C.L. and H.J. Allison 1961. Bibliography of Fossil Vertebrates, 1949-1953. Geological Society of America Memoir, Vol 84
Chang & Smith 1992
Daeschler, E.B., N.H. Shubin, and F.A. Jenkins 2006. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440:757-63
Janvier 1996
Jarvik, E. 1980
Jarvik, E 1996 . "The Devonian tetrapod Ichthyostega". Fossils & Strata 40: 1–213.)
Laurin et al. 2007
Long 1995
Schultze and Trueb 1991
Glossary