Athena Review Vol. 5, no. 1
Records of Life: Fossils as Original Sources
1. Introduction, and Geological Periods
For those who believe that the natural tendency of everything is to run down, to fall apart, and be reduced to random disorder, due to entropy -- consider a four letter word: life. It is true that living organisms, just as inanimate objects such as rocks, or human-made machines, are made up of chemicals and molecules, which themselves are subject to decay or depletion. Life, however, means reproduction and continuance. Even the simplest of living organisms (one-celled bacteria) by about 3.5 billion years ago had internal organization that accumulated and stored energy, coordinated metabolism, and programed reproduction. Photosynthesis, a complex biochemical process that is the foundation of all carbon-based life and the source of oxygen in our atmosphere, developed before 3.5 billion years ago, and will continue as long as there is sunlight and water available.
As geologist Preston Cloud (1911-1991) once wrote: "The simplest organisms were already more complex than the most complicated physical systems we know. ... The oldest possible fossil remains, however, have the sizes and shapes of bacteria. … Such wee creatures may not look very clever, but they are extraordinarily inventive biochemically. They have explored every conceivable habitat, and some inconceivable ones …And one may be sure that their ancestors were a match for the monotonously anoxic habitats of Archaean history. Bacteria even evolved the ability to repair molecular dislocations in their DNA caused by high energy ultaviolet light..." (Cloud 1989).
The Precambrian (lasting four billion years, from 4.5 to 0.5 billion years ago) was almost entirely populated by the very tiny, bacteria-like organisms described by Prof. Cloud. The records of this earliest, yet very persistant microbial life have been picked up only during the last half century, from both fossils and very sophisticated techniques of geology, biochemisty, and microscopy. This discovery process could only have happened after the development of high-magnification microscopes, photography, radioisotope dating methods, computers, and various biochemical techniques. These, except for early microscopes, didn’t exist in the late 18th and early 19th centuries, when all the other, later periods – from the Cambrian through the present- were first being discovered and described.
The entire subject of descriptive natural history was revolutionized by the invention of the microscope in about 1600, which initiated the science of biology and, in particular, histology, or the study of cell structures. During this phase (about 1600-1850), skilled drawings or renderings such as those by Robert Hooke (1680) and Anton von Lieuwenhoek (1666) were essential to reporting their microscopic discoveries of cells, bacteria, and other essential elements of biology.
Further innovations in technology, such as photography from the mid 19th century, the electron microscope from the mid 20th century, and the discovery of radioactive dating methods of ancient rock formations, have substantially raised the standards of accuracy and comparability of the records of life.
Fossils as ancient life forms.
The initial realizations that fossils actually represent ancient life forms came readily enough to some shrewd observers, such as Shen Kuo during the 11th century AD Song Dynasty in China, or Leonardo da Vinci during the 15th-16th century Italian Renaissance, but only gradually for many others. The failure to grasp this about fossils has been due to a number of preconceptions, such as about the age of the Earth. The realization that fossils were ancient plants or animals, meanwhile, effectively launches the subject of paleontology, the study of fossils.
Most of us are familiar with fossil shells and ferns (usually seen as imprints), and the reconstructed skeletons of dinosaurs and mastadons seen in museums, all typical of a series of geological periods. The study of fossils is intimately tied up with geology, the study of the earth and its rock formations. Both the discovery of the fossils themselves, and the identification of them with geological periods, were pioneering tasks involving many individuals from the 16th century onward, some of whom have also more recently discovered ways of dating the rocks containing the fossils, mainly by recording the half-life of radioactive isotopes of basalts and other igneous rocks.
The concept of huge stretches of time was also learned only gradually, the main impetus for this coming from geology. This concept of deep geological time came into sharp focus in the early 19th century with the writings of Lyell (1833), Murchison (1835), and others who had studied superimposed layers of fossils, each tens or hundreds of meters thick. All of this directly inspired Darwin (1859), who applied the uniformitarinism of Lyell (that great changes occur slowly over great amounts of time) to the evolution of animals and plants. While huge gains were made in defining geological periods and their fossil indicators in the 19th century by pioneer paleontologists ranging from Cuvier and Agassiz to "Strata" Smith (described in Section 3), it was well into the mid-20th century before anything definite was known about life before the Cambrian period - the immense span of Precambrian time, covering about 4 billion years - or even that the Earth was that old.
Whether the development of complex life on Earth from the much earlier, Precambrian time of cyanobacteria was gradual, with step-by-step accumulations of changes, simply not yet visible in the fossil record, or not preserved-- a scheme that Darwin and Lyell believed, and as many paleontologists also believe today (cf. Conway Morris 1999)-- or happened in leaps and spurts (cf. Eldridge and Gould 1977, and others), remains a subject of lively debate. The fossil record, always subject to chance preservation, and sampling error, definitely shows a sudden appearance of very complicated life forms in the time range of 540-500 million years ago (mya), during the so-called “Cambrian explosion.” The variety of multi-celled animals that first appear in the Cambrian include the earliest vertebrates or notochords (precursors of primitive fish), and a whole cast of marine invertebrates, ranging from sponges and jellyfish to very bizarre-looking crustaceans and arthropods (some worthy of the storybook creations of a Dr Suess). This begins the process of development leading from the earliest fish-like vertebrates, to mammals and to ourselves, Homo sapiens.
The subject of vertebrate paleontology (cf. Benton 2005), depends on a far-flung network of information from many sources and methods of recording, describing, and analyzing fossil data: from geology, biology, genetics, chemistry, paleontology, and the science of classification or taxonomy- in every possible combination. If the 17th century microscope revolutionized the description of biological forms by revealing the many miniature structures of life, the massive accumulation since then of data on fossil species, plus the recording of genetic sequences since the late 20th century, have enabled a rewriting of the whole structure of phylogeny or systematics- the arranging of life into related ancestral or family groups. While there are many surprises (and stumbling blocks) in the workings of paleontology, it is all perhaps made a bit more comprehensible by an awareness of the great age of the earth and its many life forms.
Geological Periods.
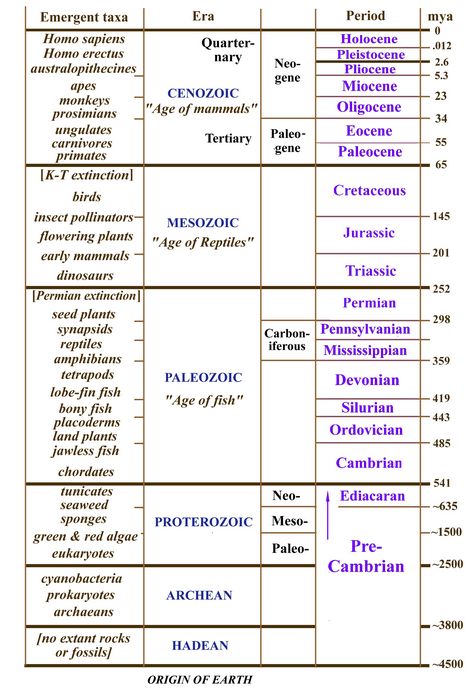
One set of terms desirable for the reader on paleontology to learn are the names and approximate dates of the geological periods. There are 18 periods (fig.1) of which the earliest, and by far the longest, is the Precambrian. Most of the period names were devised in the 19th century, an era of major advancements in the understanding of geological strata recording the history and structure of the earth, as well as the fossil evidence that went with each period. As mentioned in Section 3 on Stratigraphy, the present system of geological periods was founded on work by Smith, Sedgwick, Murchison, Lyell, and others between 1800 and 1850, in a successful application of geological stratigraphy to paleontology in the British Isles,
 Europe, and Russia.
Europe, and Russia. The Precambrian (4.5 billion to 541 mya) was not even sketchily known until the mid-20th century, although its existence was suspected by geologists in the early 19th century (Schopf 1999; Knoll 2004). As further discussed in Section 6, the Precambrian is eight times as long as the remaining seventeen geological periods put together. The Precambrian lasted from the creation of the earth (estimated at about 4.5 billion years ago) to the beginning of the Cambrian period (541-485 million years ago), when complex marine animals began to flourish. For most of these 4 billion years, the only living inhabitants of Earth were one-celled microbes resembling bacteria. Some of these, called Archaea, were able to survive in extremely hostile environments (Woese 1987; Cloud 1988; Margulis and Schwartz 1988).
The next seven geological periods, Cambrian through Permian, lasting from 541-252 million years ago (mya), comprise the Paleozoic, or "ancient life" era. This time span of 292 million years included the initial development of vertebrates from chordates through fishes, tetrapods, amphibians, reptiles, and the synapsids, who ultimately became the ancestors of mammals.
Fig.1: Timescale of geological periods, with listing of emergent lifeforms by era.
Paleozoic Periods.
The Cambrian period (541-485 mya) is marked by a rapid, substantial increase in the complexity of multi-celled organisms, including a number of marine orders such as crustaceans, mollusks, and bivalves. Since the known range of fossil taxa from the Ediacaran or Vendian stage (635-541 mya) is quite limited, the transition to the very diverse fossil record in the Cambrian appears to be quite sudden. This burst of evidence of complex life forms, which begins the Paleozoic era, is usually called "the Cambrian explosion." The Cambrian has the earliest fossil records of a wide range of complex marine invertebrates, ranging from sponges, crustaceans, and arthropods to early notochords, and the first primitive fishes, the latter known from Early and Middle Cambrian shale formations in China and British Columbia, dated 535-520 mya.
The Cambrian was named in 1832 by the British geologist Adam Sedgwick, for the Roman name "Cambria" for Wales. Sedgwick, then a professor of geology at Cambridge University, had among his students the young Charles Darwin, studying the fundamentals of stratigraphy. Sedgwick's explorations of the lowest fossil-bearing rock strata in England, Scotland, and Wales eventually led him to identify dozens of trilobites, brachiopods, and other aquatic taxa, which by the 1850s he had identified as Cambrian fauna.
The trilobites and other marine fossils found by Sedgwick and his colleage Roderick Murchison were the first fossils to be identified as “paleozoic”, meaning the oldest known life forms. In 1835 the two men presented a joint paper, titled "On the Silurian and Cambrian Systems, Exhibiting the Order in which the Older Sedimentary Strata Succeed each other in England and Wales" (Murchison and Sedgwick 1835), a detailed study of stratigraphy which formed one of the starting points of the modern geological time scale.
During the Cambrian period, warm, shallow seas flanked several continents created during the breakup of a supercontinent called Pannotia, which during the Ediacaran stage of the Precambrian, had been located mainly in the southern hemisphere. Innumerable marine creatures lived in these shallow water environments,which have now turned into layers of shale containing their fossils. Land areas at this time still lacked vegetation, as green algae (Chlorphyta) had not yet developed into land plants. There were as yet no land animals, except for insects, although trace fossils on coastal rocks have shown that some marine arthropods and mollusks did occasionally also venture onto land. Coastlines, however, were still relatively barren, supporting only near-shore mats of bacteria, and further inland, only patches of lichens and fungi.
The great array of marine invertibrates which appear for the first time in the fossil record during the Cambrian period include arthropods, some resembling horseshoe crabs, and many other now-extinct forms such as the multi-legged Hallucinogenia, named for its undoubtedly bizarre appearance. There were also a large variety of crustaceans, as well as tunicates, or hemichordates, mollusks, brachiopods ("shoulder-legs", a common early type of bivalve), seaworms,sponges, and jellyfish.
Probably the most common arthropods were trilobites ("three lobes"), which are extremely diverse in form, and probably among the best studied fossils because of the relative ease of identifying and collecting them. Many of the trilobite variations were related to defenses from predators who hunted them. Among the most formidable was Anomalocaris, a large, multi-lobed arthopod up to 70 cm (2 feet) long with a large head, considered the dominant predator of Cambrian waters. It had excellent vision, being provided with a pair of large, compound eyes on stalks with nearly 16,000 individual lenses. Its disk-like mouth, used for crushing prey, was composed of 32 overlapping plates (Whittington and Briggs 1986).
All these are documented in extremely well-preserved fossil beds located in in fine-grained sedimentary rock, such as the Burroughs shale of British Columbia, first discovered by Charles Walcott of the Smithsonian Institute in 1909. More recently, extensive fossil beds in the Maotianshan shales near Kunming, in Yunnan province of southern China, have yielded a vast array of Cambrian fossils, in some cases with even better preservation than those from the Burroughs shale.
The Cambrian also saw the first appearance of chordates or primitive vertebrates. These took the form of notochords (animals with simple neural cords made of cartilege), such as the 2 cm.-long Haikouella and Yunnanozoon from the Maotianshan shales (535-520 mya). These early notochord fossils are described in more detail in Section 7, covering the development of fish from chordates.
Some are comparable to Pikaia gracilens from the Burgess shale in British Columbia (520 mya). Pikaia, found and first identifed by Walcott (1912) as a polychaete or sea worm, was much later restudied and identified as a chordate by Simon Conway Morris (1979), who later recognized similarities with the Chinese Yunnanzoon (Morris and Caron 2012). Pikaia are 1-2 inches in length and resemble a lancelet. These early chordates had gill slits and flagella or tendrils, but lacked lower jaws or teeth.
More advanced early vertebrate fossils from the Cambrian period have also been found in the Chenjiang shales. These include Haikouicthys and Myllokunmingia, thought to be the earliest fish yet discovered.
The Ordovician period (485-443 mya) was named in 1879 for the Ordovices, a Celtic tribe in Wales, by Charles Lapworth. He proposed the period as a needed transition between the Cambrian and Silurian, as then defined by Sedgwick and Murchison.
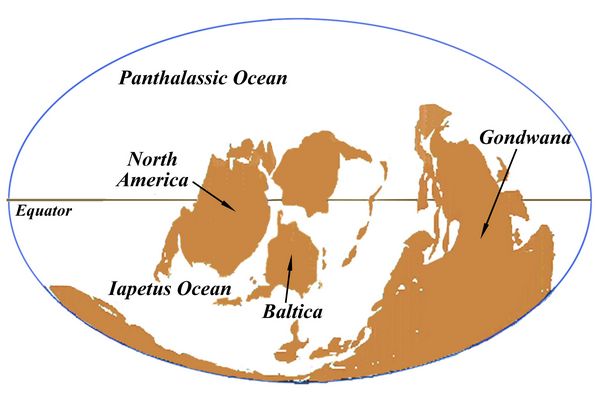
During the Ordovician, southern landmasses including what is now Africa, South America, Australia, and Antarctica were joined in the supercontinent named Gondwana, then located near the South Pole. Western and Central Europe were at that point in the southern tropics. North America, located in the Iapetus ocean around the Equator, was converging into the proto-northern Europe region called Baltica (fig.2). Their juncture would create the Taconic orogeny, causing mountain ranges to emerge. Their remnants today are source areas for Ordovician fossils, located in north-eastern North America, Greenland, Norway, and the British isles (Palaios 2014f).
The Ordovician notably saw the development of jawless fishes (agnathans) from the small, lancelet-like forms of the Cambrian into much larger, armored fish, collectively called Ostracoderms ("bony skin"). Among the most successful branches was the superclass named Pteraspidomorphi ("wing shield forms"), which lived from the Ordovician through Devonian periods.These included the orders of astrapida ("star shields"), such as Astraspis desiderata from about 450 mya, which had a lateral line sensing system and a series of eight gill openings; and the order Heterostraci ("different scales"), whose species were covered with both armor plates and scales.
Fig.2: Globe during the Late Ordovician, ca. 450 mya.
Other forms of marine animals included swimming nautiloids with well-developed eyes, who became dominant predators. Trilobites, who were hunted by them, evolved various defenses including elaborate spines and nodules. Some trilobites developed prominent eyes to help see predators. A well-known example is Asaphus kowaleski, a species with long eyestalks, found in Middle Ordovician layers on the Volkhov River near St Petersburg, Russia .
In the Early Ordovician, tabulate corals appeared, along with planktonic graptolites, conodonts, and sponge-like bryozoans. In the mid-Ordovician, rugose coral first developed, which together with tabulate corals, are the source of extensive carbonate deposits from the period. The period also saw a wide diversification in brachiopods (early bivalves) and mollusks.
The first evidence of land plants also appeared in the Ordovician. Land plants evolved from a type of green algae or seaweed called chlorophyta, similar to today's sea moss or sea lettuce, which colonized freshwater areas around the shore. Land plants at first retained the simple thalloid cell structure of seaweed (algae), and grew in tiny non-vascular forms resembling liverworts. They reproduced by spore dispersion; fossil spores from land plants have been identified in the very latest Ordovician sediments.
The Silurian period (443-419 mya) is named after the Silures, the Roman name for an ancient tribe in Wales. The stratigraphy and index fossils in Britain for this period were defined in the early 1830s by the Scottish geologist Roderick Murchison, who named the period in 1833. Murchison was a a colleague of both Charles Lyell and Adam Sedgwick.
In 1831, Murchison made an initial report to the British Scientific Association on the stratigraphy of paleozoic rock layers in the Welsh borderlands. These included (from top to bottom) Carboniferous limestone with coal-bearing strata (divided in the Americas into the Mississippian and Pennsylvanian periods) dating about 360-290 mya; Devonian “Old Red” sandstone (412-360 mya), and fault zones of schist, slate, and other metamorphic rocks identified by Murchison as “Transition Rocks”, which intruded downward into a fourth, as yet poorly defined region of sedimentary rocks, which still in 1831 had no identified fossils. This lowest, unresolved strata, today identified as a series of Silurian, Ordovician, and Cambrian layers, was thus at first designated as “agnotozoic” ("without known life").
Two years later in 1833, however, Murchison reported to the Geological Society of London that he had identified, in this lowest strata, several superimposed layers of sedimentary rocks with distinct fossils. These constituted the Silurian layers, all located beneath the previously identifed zone of metamorphic or “Transition” rocks
The layers as described by Murchison, in the the first detailed definition of the Silurian period (443-419 mya), made up a series of six formations. These included (from top to bottom): a) the Upper Ludlow rocks, now a marker for the Upper Silurian, at 427-423 mya; b) the Wenlock Limestone, now dated at 433-427 mya; c) the Lower Ludlow rocks; d) the Shelly Sandstones; e) a black shale which Murchison called “Black trilobite flagstone,” containing trilobite fossils of a diagnostic species, Asaphus buchii, found only in that layer (now renamed Ogygia bucchi); and f) a vast, still undefined lowest zone of red conglomerate sandstone and slaty schist “several thousand feet in thickness.”
As revealed by the fossils first organized by Murchison, the Silurian included significant developments in both marine invertabates and in fish. Notably, this included the first appearance of armored, jawed fish called placoderms. Recent examples from Yunnan fossil beds include several finely preserved, newly defined taxa dating from 420-415 mya. These include Sirolupis, considered to have evolved from one of the ostracoderms, and the recently discovered Entelognathus priomordialis, a placordern showing the first appearance of the jawbones (premaxilla, maxilla, and dentary) later found in bony fish, tetrapods, and all land vertebrates.
The Silurian also saw a great abundance of mollusks, brachiopods, cephalopods (squid-like animals), corals and other colonial animals such as bryozoa, and crinoids, plant-like animals attached by a stem to the sea floor, which are a characteristic fossil of the period. During the Silurian there was also significant evolution of land plants, including the first plants with specialized, vascular tissue cells.
The Devonian period (419-359 mya), named for fossils and rock formations found in Devon in southern England, was defined by Roderick Murchison and Adam Sedwick between 1830-1840. In their analyses of the stratigraphy of the British Isles, Murchison and Sedgwick identified the Devonian period with red and brown oxidized sedimentary deposits known as the Old Red Sandstone, with outcrops in the county of Devon providing type locations. Between 1834-1840 there was controversy on the definition of this period between Murchison and Henry De la Beche, first director of the Geological Survey in England, regarding the absence of the Old Red sandstone in parts of North Devon between Silurian and Carboniferous levels. At first the absence of this red sandstone, which Murchison and Sedgewick saw as marking the intervening Devonian period, prevented its clear identification. In 1840, however, Murchison discovered a layer in Russia similar to the one found in Devon, placed between well-defined Silurian and Carboniferous deposits. This conclusively supported Murchison and Sedgwick's definition of the Devonian.
Lasting from 419-359 million years ago, the Devonian period saw major shifting of continental plates related to undersea tectonic and volcanic action. Convergence of the continents of North American and Baltica in the Early D
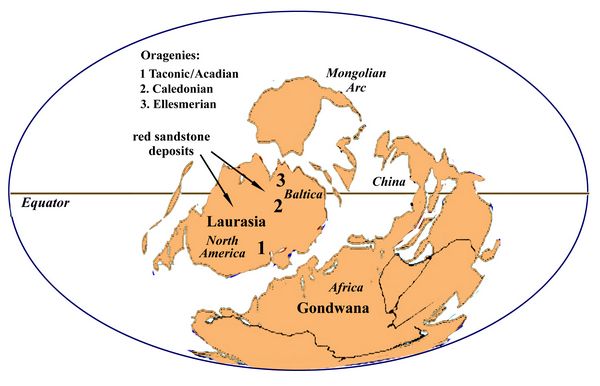 evonian created the continent called
Laurasia, which contained large zones of arid, near desert conditions
(fig.3). This is the source of the Old Red Sandstone beds, which
contain hematite, typical of drought conditions. The collision of
continental plates also caused the Taconic and Caledonian orgenies,
resulting in the uplift of the northern Appalachian mountains of New
York state and Canada, and the Caledonian mountains in the British
Isles and Scandanavia (fig.3). By the Permian period, Laurasia and
Gondwana would merge into the supercontinent Pangaea (fig.4).
evonian created the continent called
Laurasia, which contained large zones of arid, near desert conditions
(fig.3). This is the source of the Old Red Sandstone beds, which
contain hematite, typical of drought conditions. The collision of
continental plates also caused the Taconic and Caledonian orgenies,
resulting in the uplift of the northern Appalachian mountains of New
York state and Canada, and the Caledonian mountains in the British
Isles and Scandanavia (fig.3). By the Permian period, Laurasia and
Gondwana would merge into the supercontinent Pangaea (fig.4).Fig.3: Globe during the Early Devonian, ca. 400 mya.
High sea levels prevailed during the Devonian, known as the "Age of Fishes". Marine invertebrates were dominated by brachiopods, bryozoa, corals, crinoids (which resembled lilies but were animals), and trilobites, which still remained common. Among vertebrates, the armored agnathans or jawless fish known as ostracoderms declined, as many branches of jawed fish (gnathostomes) expanded. Armored placoderms were numerous during the lower stages of the Devonian Period, but became extinct in the Late Devonian One of the latest widespread placoderms was the antiarch Bothriolepis canadensis, found first near Quebec, Canada, from about 380 mya. Also appearing were early cartilaginous fish (Chondrichthyes), including the widespread shark Cladoselache.
The Devonian saw dramatic developments in the later stages, when tetrapods first appeared. This was preceded by a branching of bony fishes (Osteichthyes) into two main orders, ray-finned fishes (actiopterygians) and lobe-finned fishes (sarcopterygians). The pectoral fins of the latter were modified into a pair of long, arthropod-like limbs which had elbow-like joints. These were probably used originally for burying and raising themselves out of the mud substrate. They led, however, to the evolution of tetrapods, still fish-like, but the direct ancestors of amphibians.
Tetrapods first appearred during the Late Devonian. (380-360 mya) Basically they began as lobe-finned fish (sarcopterygians), who became adapted to fresh water inlets. They developed lungs to breath oxygen out of water, and then colonized mudflats and shorelines. Examples are Acanthostega gunnari, Pederpes, and Eusthenopteron fordii. This sequence, for which there is increasingly detailed fossil evidence (Clack 2012), is discussed in the sections 10 and 11 on sarcopterygians and tetrapods. The Devonian also saw a major expansion in land plants, including the development of more complex reproductive cycles.
There was then a great radiation of large amphibians iduring the Mississippian and Pennsylvanian periods (359-298 mya). These two periods together are called the Carboniferous in the Eastern Hemisphere. By the Pennsylvanian period (328-298 mya), some branches of amphibians developed into reptile-like amphibians called Sauropsids, that are ancestral to both reptiles and synapsids. Both large and small reptiles and synapsids then expanded, at the expense of amphibians, throughout the drier Permian period (298-252 mya), until a huge extinction event at the Permian/Triassic boundary eliminated an estimated 90% of all vertebrate species.
The Mississippian period (359-323 mya), named for marine limestone rock exposures found in the Mississippi valley, lasted from 360-325 million years ago. In North America, the Mississippian limestone beds and the subsequent Pennsylvanian, with its extensive coal deposits, have been treated as separate geological periods. During the Mississippian, plate tectonics and orogeny (collision of continental plates) created the Appalachian Mountains. It was a relatively warm period with an expansion of ferns and coniferous forests, whose legacy included vast coal fields and petroleum deposits.
In Europe, the equivalant periods of the Mississippian and Pennsylvanian shows a more continuous sequence of lowland continental deposits and are grouped together as subdivisions of the Carboniferous Period. In the Northern regions most of the land areas were then transgressed by shallow seas, resulting in extensive carbonate deposits now turned into marine imestones. Only higher areas such as the Fennoscandian and Laurentian Shields remained above sea level, surrounded by river delta systems and lagoons.
The Mississippian saw a radiation of land vertebrates, with a great diversification of amphibians, and the first appearance of Reptilomorphs, or reptile-like amphibians.
The Pennsylvanian period (323-298 mya), the second of the Carboniferous periods in the Western hemisphere, is named for the vast Carboniferous (coal) deposits in the hills of western Pennsylvania. Coal, found in organic-rich beds within layers of shale and sandstone, contains many fossil imprints, especially of plants.
The Pennsylvanian is correlated with four Russian stages, from oldest to youngest, Bashkirian, Moscovian, Kasimovian, and Gzhelian. The collapse of extensive rainforest environments to cooler, drier conditions, in the mid Pennsylvanian (between the Moscovian and the Kasimovian) eliminated amphibians who could not adapt to cooler, drier conditions. Those who did adapt included larger amphibians such as Dendrepeton, who inhabited forest wetlands and grew up to several meters long.
A key evolutionary innovation in the context of a drier environment was the amniotic egg, encased in a waterproof membrane which did not need to be immersed in water. This allowed far wider exploitation of the land by some branches of small, lizard-like tetrapods. The amniotic egg is considered to define the earliest sauropsid reptiles including Hylonomus, an insectivore dating from 312 mya, and the earliest known synapsid, Archaeothyrus a larger, ground-dwelling carnivore about a half meter in length, dating from 306 mya. Fossils of both, as well as of Dendrepeton, have been found in Joggins, Nova Scotia, then a subtropical swampland filled with Lepidodendron and other huge plants related to club mosses.
Sauropod reptiles and synapsids underwent a major evolutionary radiation, expanding in many areas in response to the drier climate that followed the rainforest collapse.
The Permian period (298-252 mya) was defined in 1841 by the Scottish geologist Roderick Murchison (already instrumental in defining the Silurian and Devonian periods), who explored fossil zones in the Ural region of western Russia, and named the period for the ancient kingdom of Permia, based in today’s region of Perm. Murchison had first recognized in 1840 these red-bed formations, which he explored between St. Petersburg and Moscow (Benton 2005). This part of Russia, like the oil-rich Permian basins in Texas and Oklahoma, has numerous deposits containing vertebrate fossils exposed at or near the surface of frequently cross-bedded shales and sandstones, which represent ancient river courses and lakes.
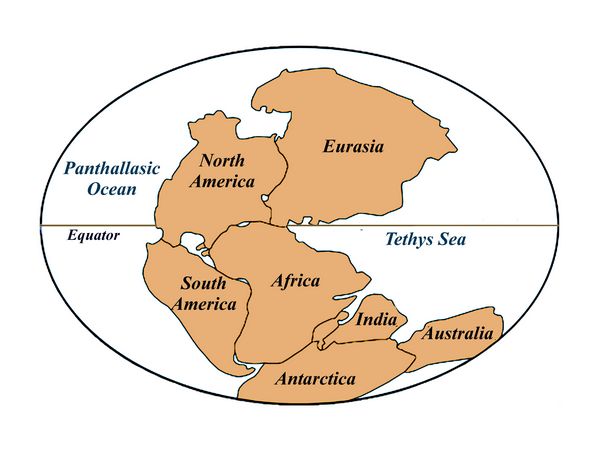 This period, the last of the Paleozoic era,coincides with the formation of
Pangaea, the supercontinent
with all major land masses except
eastern Asia (fig.4). The extensive rainforests of
the Mississippian and Pennsylvanian periods by now had vanished,
replaced
by vast inland desert regions on the continents.
This period, the last of the Paleozoic era,coincides with the formation of
Pangaea, the supercontinent
with all major land masses except
eastern Asia (fig.4). The extensive rainforests of
the Mississippian and Pennsylvanian periods by now had vanished,
replaced
by vast inland desert regions on the continents.Reptiles, whose amniotic reproduction allowed them to adapt to such dryer conditions, became dominant, replacing their amphibian ancestors. Major expansions of reptile groups occurred including the ancestral groups of mammals (synapsids), turtles (anapsids), and archosaurs (diapsids) who were the ancestors of dinosaurs, crocodiles, and birds.
Fig.4: Globe during the Permian showing Pangaea.
Both the Permian Period and the Paleozoic era ended with the largest known mass extinction on Earth, in which it is estimated that nearly 90% of marine species and 70% of terrestrial species died out. It would take well into the Triassic for life to recover from this catastrophe.
Mesozoic Periods: Triassic, Jurassic, and Cretaceous
The first small mammals appear in the next two periods, in the Late Triassic and the Early Jurassic (about 220-180 mya). These two periods, along with the subsequent Cretaceous, comprise the Mesozoic ("middle life") era, from 252-65 mya. This era, also known as the age of reptiles, saw the rise and fall of the dinosaurs, and ended with another major extinction event at the the "K-T boundary". [In this phrase, K is the geologic map symbol for the Cretaceous period; while T stands for "Tertiary", an older name for the next five periods, that make up the Cenezoic era (the era of "recent life").] Note that both the Paleozoic and Mesozoic eras ended with big extinction events.
Cenezoic Periods.
These five Cenezoic periods, lasting from 65 to 2 mya (all ending in -cene ["recent"]), are the Paleocene, Eocene, Oligocene, Miocene, and Pliocene. Worldwide, the Cenezoic era is filled with all types of mammals expanding into new habitats from which large reptiles have now mostly disappeared, and is called the Age of mammals. By the end of this era, the earliest ancestors of humans have developed from earlier, ape-like primates. Then comes the period of intermittant glaciers called the Pleistocene, from 2 million to 12,000 years ago, which saw the whole span of the evolution of Homo (our genus). Following that comes our own brief period, called the Holocene ("fully recent") period.
References:
Benton 2005
Clack, J.A. 2012. Gaining Ground
Cloud, P. 1989. pp. 42, and 233-234.
Conway Morris, Simon 1979
Conway Morris and Caron 2012.
Eldridge and Gould 1977
Knoll, A. 2004
Lapworth
Lyell, C. 1833
Margulis and Schwartz 1988
Murchison, R.I. 1831
Murchison, R.I. and A. Sedgwick 1835. On the Silurian and Cambrian Systems, Exhibiting the Order in which the Older Sedimentary Strata Succeed each other in England and Wales.
Murchison, R.I. et al. 1845
Palaios 2014f.
Schopf 1967, 1999
Walcott, C. 1912
Whittington and D. Briggs 1986.
Woese C. R. 1987. "Bacterial evolution." Microbiology Reviews 51, pp. 221–271.
Glossary