|
Michele A. Miller, PhD Editor of Old World Archaeology, Athena Review
.
The fate of Neanderthals has always been hotly debated, beginning with the
discovery of their first remains in the 19th century
(see timeline).
Discussions about Neanderthal existence and their relation to modern humans
have at times been vehement, often leading to personal rivalry not only in
the scholarly community,
but also within the realm of popular culture. Yet what is it about these
robust and stained old bones that has lead to so much discussion and dispute?
In part, the answer is that the way we view Neanderthals reflects how we
view ourselves, as Neanderthals are the closest known relatives to anatomically
modern humans. Are we truly the unique and especially blessed creations of
God, or merely the latest branch in a tree which once blossomed with many
other similar life-forms?
The last 150 years have seen continuous debate in which Neanderthals have
been either dismissed as mere dead-end, distant cousins of 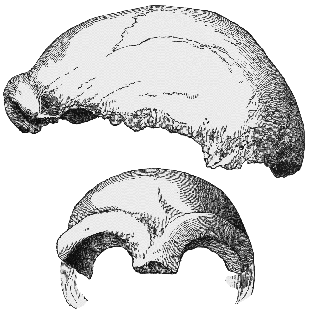 modern Homo sapiens,
or embraced as our direct ancestors. By the late 18th century, an age when
many believed the Bible recounted the only true early history of humankind,
recognition of extinct species, although only accepted as
“ante-diluvial,” or destroyed in the Biblical flood, was a
revolutionary step by Cuvier, Buffon, and other early paleontologists. The
19th century discovery of fossilized Neanderthal bones was, therefore, essential
in the early debate over the antiquity of man and the ultimate acceptance
of evolutionary theory. modern Homo sapiens,
or embraced as our direct ancestors. By the late 18th century, an age when
many believed the Bible recounted the only true early history of humankind,
recognition of extinct species, although only accepted as
“ante-diluvial,” or destroyed in the Biblical flood, was a
revolutionary step by Cuvier, Buffon, and other early paleontologists. The
19th century discovery of fossilized Neanderthal bones was, therefore, essential
in the early debate over the antiquity of man and the ultimate acceptance
of evolutionary theory.
Fig.1: Huxley's drawing of the original Neanderthal fossil found in
the Neander Valley in 1856, showing the low but large skull, and accentuated
brow ridges (T.H. Huxley, 1863).
The first Neanderthal remains, although not recognized as such for several
decades, were found in 1830 in Engis Cave, Belgium and in 1848 at Gibraltar.
A few years later, fossils found at Feldhofer Cave in the Neander Valley
of Germany (fig.1) touched off considerable debate. While a very few championed
the remains as the ancient species of “Neanderthal Man,” many others
rejected them as merely the pathological skeleton of a deformed individual.
When Darwin published his theory of natural selection in 1859, he avoided
applying the mechanism of evolution directly to humans, ending his manuscript
only with the enigmatic words, “and light will be thrown on the origin
of man and his history.” While Darwin did not yet press the issue, others
were quick to see the implications of his work. Only a few years after the
initial publication of On the Origin of Species, Thomas Henry Huxley,
Darwin’s most prominent supporter, published the first scientific assessment
of the Neanderthal and Engis fossils (fig.1).
The extinct human
species, Homo neanderthalensis, was first proposed in a controversial paper
read in 1864 by William King, a professor from Queen’s College in Ireland.
King thought the Neanderthal skull “more closely conforms to the braincase
of a chimpanzee than a human.”
King’s consignment of Neanderthals to a different species from humans
was based o n a comparison with hunting and gathering people from the Andaman
Islands near India, whose physical traits, “viewed in connection with
the strictly human conformation of his cranium, ...specificially identify
him with Homo sapiens. Psychical endowments of a lower grade than those of
the Andamaner cannot be conceived to exist; they stand next to brute
benightedness.” (in Trinkaus and Shipman 1993, pp.88-89). T.H. Huxley,
by contrast, on purely anatomical grounds compared two Neanderthal skulls
with those of Scandanavians and Australian aborigines, and found Neanderthals
to be within the human range. n a comparison with hunting and gathering people from the Andaman
Islands near India, whose physical traits, “viewed in connection with
the strictly human conformation of his cranium, ...specificially identify
him with Homo sapiens. Psychical endowments of a lower grade than those of
the Andamaner cannot be conceived to exist; they stand next to brute
benightedness.” (in Trinkaus and Shipman 1993, pp.88-89). T.H. Huxley,
by contrast, on purely anatomical grounds compared two Neanderthal skulls
with those of Scandanavians and Australian aborigines, and found Neanderthals
to be within the human range.
Fig.2: Neanderthal skullcap (calotte) from Spy, Belgium showing
characteristic brow ridges (Athena Review, from cast at AMNH).
As the theory of evolution gained acceptance, so did the recognition of
Neanderthals as ancient, anatomically-different, humans. By 1866, the Neanderthal
jaw discovered at Trou de la Naulette, Belgium, was declared, by the anatomist
Paul Broca, to constitute “the first fact that furnishes anatomical
evidence to the Darwinists. It is the first link on a chain that extends
from man to the apes” (Trinkhaus and Shipman 1993, p.103).
While it came
to be accepted that Neanderthals were an ancient form of human, what remained
to be determined, and what is still hotly debated, is the exact relationship
of these robust and (as many consider) “primitive”-looking beings
with modern humans. By the early 1900’s, scholars like Gustav Schwalbe
and Aleš Hrdlicka believed Neanderthals were direct ancestors of modern
humans, while others like Arthur Keith and Marcellin Boule suggested Neanderthals
were an evolutionary side-branch, an extinct species that had nothing to
do with human ancestry. These same evolutionary ideologies still exist today,
in the form of the multi-regional continuity vs. replacement theories.
In part, where Neanderthals were placed on the human evolutionary tree was
largely influenced by how their anatomy and behavior were viewed. Boule,
for instance, who studied the Neanderthal remains from La Chapelle-aux-Saints
(fig.3) and La Ferassie, described Neanderthals (based on several incorrect
anatomical analyses) as stooping in posture, with a slouching, b ent-kneed
gait-altogether too apish to be ancestral to modern humans. Boule sought
to distance modern humans from what he saw as brutish beings, arguing forcefully
that the Neanderthals had been annihilated by the more
“elegant…inventive” Homo sapiens, a scene retold in various
20th century novels such as William Golding’s The Inheritors. ent-kneed
gait-altogether too apish to be ancestral to modern humans. Boule sought
to distance modern humans from what he saw as brutish beings, arguing forcefully
that the Neanderthals had been annihilated by the more
“elegant…inventive” Homo sapiens, a scene retold in various
20th century novels such as William Golding’s The Inheritors.
Fig.3: Neanderthal skull from La Chapelle-aux-Saints in France
(Athena Review, from cast at AMNH).
In a similar way, Karl Gorjanovic-Kramberger, the Croatian paleontologist
who excavated and analyzed the large sample of Neanderthals from Krapina,
distanced modern humans from Neanderthals by proposing fragmented skeletons
from the site provided evidence of cannibalism. “These men ate their
fellow tribesmen, and what’s more they cracked open the hollow bones
and sucked out the marrow,” (Pracovjek iz Krapine, 1918). The specter
of the Neanderthal as a cannibal survives today in the recent research at
Moula-Guercy Cave in France by Defleur et al. (1999).
These interpretations of Neanderthal behavior fed neatly into a prevalent
“pre-sapien” theory of human origins proposed by Boule and Keith,
who believed the human lineage had split at some remote time in the past
(ie., during the Pliocene, 5-2 mya) into two separate branches. One of these
branches led to modern humans, while the other ended in extinct species,
including Neanderthals. A general belief in this theory may have helped many
otherwise astute scientists accept the authenticity of the infamous Piltdown
skull in Kent, England
(timeline, phase 3).
It was far more comfortable to consider our ancestors to have large modern-like
brains, if a rather ape-like jaw, then to look for human origins in the
primitive-looking (if large-brained) skulls of Neanderthals or the much older,
smaller-brained fossil hominids uncovered between 1924-1927 in Africa
(Australopithecus; fig.4) and China (Homo erectus).
Surprisingly, it was another claim of Neanderthal cannibalism that actually
lead to the view that Neanderthals were more human-like than previously believed.
Alberto Blanc interpretated a skull from Monte Circeo, Italy in 1939 as
indicating cannibalistic but also mortuary practices. Along with an enlarged
opening at the base of the braincase, which Blanc claimed occurred as the
brain was extracted for a feast, he also proposed that the skull was found
buried in a circle of stones. While Blanc's evidence for Neanderthal ritual
was later questioned, his theories lead to renewed belief in the humanity
of Neanderthals.
Even before Blanc's
discovery, various researchers had suggested that Neanderthals exhibited
ritualistic behaviors. For instance, excavations from 1917 to 1921 at Drachenloch
(“Dragon's Lake”) in the Churfirsten Mountains of Switzerland uncovered
Mousterian tools alongside numerous bones of Ursus spelaeus, the huge cave
bears that became extinct 50,000-40,000 years ago. The a mateur archaeologist
Dr. Emil Bächer described the bear bones as found within several stone
“cists” capped with stone slabs or deliberately piled up behind
heaps of stones. Although later analyses revealed the accumulations resulted
from the natural deaths of the bears during hibernation, Bächer saw
them as evidence for Neanderthal cave bear cults, in which the bears were
the subject of worship and perhaps sacrifice, an idea employed in 20th century
popular literature, including Clan of the Cave Bear by Jean Aeul. mateur archaeologist
Dr. Emil Bächer described the bear bones as found within several stone
“cists” capped with stone slabs or deliberately piled up behind
heaps of stones. Although later analyses revealed the accumulations resulted
from the natural deaths of the bears during hibernation, Bächer saw
them as evidence for Neanderthal cave bear cults, in which the bears were
the subject of worship and perhaps sacrifice, an idea employed in 20th century
popular literature, including Clan of the Cave Bear by Jean Aeul.
Fig.4: Type specimen of Australopithecus africanus with incomplete
skull and limestone endocast from Taung, South Africa, found by Raymond Dart
in 1924 (Athena Review, from cast at AMNH)
As Neanderthal behavior began to appear more human, some scholars accepted
the idea that they were our own ancestors. For instance, at this time Aleš
Hrdlicka suggested a unilinear theory of human evolution, in which Neanderthals
were merely one stage leading to Homo sapiens. The idea (perhaps
originating with T.H. Huxley) that Neanderthals were not very physically
different from modern humans eventually began to gain wider acceptance.
After World War II, some of the greatest barriers to the acceptance of
Neanderthal’s “humanity” collapsed. Thanks to fluorine dating
tests conducted by Oakley in the early 1950’s there was conclusive proof
that the troublesome Piltdown remains were a forgery consisting of an ancient
cranium of an anatomically modern human with the purposely stained and filed
jaw of an orangutan. Only a few years later, two major re-analyses of the
skeletal material from La Chapelle by C. Arambourg, and Straus and Cave,
finally put to rest the notion of the bent-kneed, bent-over, apish Neanderthals.
Moreover, Etienne Patte demonstrated the anatomical characteristics observed
in the Neanderthal fossils were statistically not much different than those
seen in the full range of modern humans. Following Coon (1939), Straus and
Cave observed that, if bathed, shaved and dressed, Neanderthals would hardly
attract attention on a New York subway. This was reinforced by Garrod’s
discovery in the 1930’s at Mount Carmel of so-called
“progressive” Neanderthals.
It was during
this phase of synthesis among possible species that Franz Weindenreich developed
the multi-regional theory of human origins that still is accepted by several
prominent researchers today. According to this theory, the regional variations
in modern human form that we call “racial” differences originated
in geographically separated populations of Homo erectus. An
“ultra-lumper,” Weindenreich believed that the various ancient
hominid fossils which had heretofore been “split” into many different
species (and even genera) should all be considered as a single species, Homo
sapiens.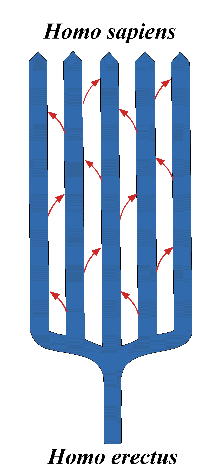
In his 1947 publication, Weidenreich proposed that a single, ancient form
of this species had long ago spread across the Old World, developing separately
to form modern humans. Although this allowed for localized features to develop
in humans in each area, Weidenreich also postulated that there was considerable
interbreeding and therefore genetic exchange between the various regional
groups, which can be better visualized as the intersecting diagonal lines
between regional groups in an evolutionary diagram (fig.5). One corollary
of his theory is that Neanderthals would be considered merely one of many
regional representatives of a particular stage of human development, with
similar ancient human stages apparent in geographic areas outside Europe.
Fig.5: "Candelabra" model of human evolution (arrows show genetic
intermixing).
In ensuing debate by 1959, William Howell challenged Weidenreich's ideas
with a “Noah’s Ark” model in which humans developed from a
single, recent origin and replaced the earlier hominids without interbreeding
or genetic exchange. Three years later, Carleton Coon postulated a highly
controversial theory for the origin of modern humans similar to
Weidenreich’s but adding that Homo erectus not only developed into Homo
sapiens separately in each region, but did so at different rates. In the
politically charged sixties, Coon’s suggestion that “Caucasoids”
(Europeans) and “Mongoloids” (Asians) had developed into Homo sapiens
before native African and Australian peoples was rejected as racist.
Many of Coon’s other ideas were better accepted, especially his proposal
that the particularities of Neanderthal anatomy reflected adaptation to the
cold, glacial climate of Pleistocene Europe. For instance, he noted that
the short-limbed stocky build of Neanderthals mirrored that of modern Eskimo
tribes, while human populations closer to the equator tended to be more slender
and longer-limbed, otherwise known as Allen’s rule. A more squat, stocky
build enables a mammal to conserve body heat since it minimizes the surface
area of the body relative to volume. Similarly, with his “radiator
theory” of Neanderthal noses, Coon noted that the Neanderthal’s
large nasal apertures and prominent nose served to warm the air as it was
inhaled, thus protecting the delicate brain stem from the cold air of their
surroundings (for recent, contrary findings, see Mowbray and Gannon this
issue).
In 1946 Earnest Hooton brought attention to the heterogeneity between
“classic” European Neanderthals and “progressive” central
European and Near Eastern Neanderthals, making interspecific comparisons
more complex.
As controversies with racial comparisons continued, exciting new discoveries
made by Ralph and Rose Solecki at Shanidar Cave in Kurdistan, Iraq added
fuel to the idea that Neanderthals were culturally more similar to ourselves
than previously acknowledged (fig. 6). Between 1953 and 1960 nine Neanderthal
skeletons were found at Shanidar, all more similar in anatomy to the
“classic” Neanderthals of Europe than the more
“progressive” type found at Tabun cave in Israel. While four of
these individuals seem to have been buried in the cave due to natural causes
(probably caught under rock fallen from the cave ceiling), the other five
appeared to have been intentionally buried. Further, the discovery of large
amounts of pollen in the soil by the skeleton of one elderly male suggested
to Solecki that the individual had been buried with offerings of flowers.
Adding to such signs of humanity in the Shanidar remains, analysis of the
bones of another male Neanderthal showed he had lived well past maturity
despite sustaining a number of injuries earlier in life , which may have
left him both blind and considerably crippled, without the use of his right
arm. In his 1971 book about the cave discoveries, tellingly titled Shanidar:
The First Flower People, Solecki pointed out that such a significantly
handicapped individual would not have survived without the care and protection
of his fellow Neanderthal community.
With Neanderthals looking more and more human in their behavioral
characteristics, scholars once again argued to place them on the human lineage.
At the same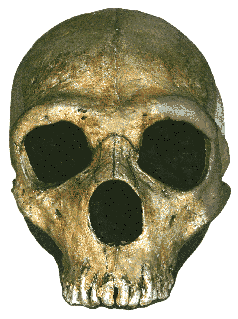 time
that Coon's theories were being debated among American anthropologists, C.
Loring Brace refocused attention back to the Neanderthal question, arguing
that Neanderthal anatomy was less highly specialized than had been formerly
proposed. Moreover, he asserted that a gradual change could be seen in
tool-making traditions from the Mousterian into the Upper Paleolithic, thus
contradicting the sudden replacement of Neanderthals by modern humans. Today
the replacement versus continuity debate remains prominent
(see Shea, this issue). time
that Coon's theories were being debated among American anthropologists, C.
Loring Brace refocused attention back to the Neanderthal question, arguing
that Neanderthal anatomy was less highly specialized than had been formerly
proposed. Moreover, he asserted that a gradual change could be seen in
tool-making traditions from the Mousterian into the Upper Paleolithic, thus
contradicting the sudden replacement of Neanderthals by modern humans. Today
the replacement versus continuity debate remains prominent
(see Shea, this issue).
Fig.6: Neanderthal cranium from Shanidar, Iraq (Athena Review,
from cast at AMNH).
In a later paper Brace claimed that most of the differences between Neanderthals
and modern humans were in the dimensions of their face and teeth, and that
this reduction in size of the face and teeth that occurred over time could
be explained as the by-product of cultural evolution. As specialized tools
replaced the use of teeth for various tasks, for instance, the size of teeth
and facial dimensions would eventually be reduced. In essence, Brace suggested
that for humans, culture was more important than biology in determining the
direction of evolutionary change. Later, Brace argued that not only was culture
the human “ecological niche” but, as according to ecological theory,
only one species can occupy a certain niche at a time, so only one human
species could occur at one time. This version of the unilineal theory of
hominid evolution soon became widely accepted as the “single species
theory” during the late 1960’s and early 1970’s. With the
fossil discoveries at the end of that decade leading to proposals of several
early hominid species (i.e., Homo ergaster, H. habilis and
H. rudolfensis) living contemporaneously in Africa, however, new
formulations were required.
By the early 1970's Milford Wolpoff had reinstated Weidenreich’s theory
of multiregional continuity as a mechanism for the evolution of modern humans.
Seeing evidence within the fossil record for continuity of certain physical
characteristics within geographic regions, Wolpoff reasoned that these specific
variations originated with Homo erectus. Unlike Coon, however, Wolpoff recognized
the key role of gene flow between otherwise isolated human populations. Without
this genetic exchange it would be impossible to propose that each population
would have evolved into the same species, Homo sapiens. This continuity of
anatomical characteristics is perhaps least obvious in the comparison of
“classic” European Neanderthal skeletal material to modern humans.
However, in an effort to prove his position, Wolpoff compared teeth from
the more transitional Krapina Neanderthals with fossil teeth from early Homo
sapiens, finding a long-term evolutionary trend for increasing size in molars
and decreasing size in incisors from former to later.
By the middle of the 1970's a new crop of scholars were adding to the debate.
The first of these was Christopher Stringer, whose 1974 dissertation focused
on the quantitative analysis of Neanderthal crania. Stringer observed too
many differences between Neanderthal and modern human crania to believe that
Neanderthals could have been our direct ancestors, and thus became a strong
proponent of the “replacement hypothesis.” After noting that several
skulls found in Ethiopia (Omo 1 and 2), dated to between 130 and 40 thousand
years ago, appeared anatomically modern, Stringer proposed that Homo sapiens
originated in Africa. Only two years later, however, Fred Smith presented
his dissertation with remarkably different conclusions that, based on a new
quantitative study of the Krapina Neanderthal material, supported the
multiregional hypothesis that Neanderthals had evolved into modern humans
within Europe.
By the middle of the decade, the opposing camps in the Neanderthal debate
were clear. Smith and Wolpoff followed in the footsteps of Brace in asserting
that Homo sapiens evolved indigenously in Europe from Neanderthals.
Stringer, on the other hand, just as adamantly declared that modern humans
evolved outside Europe, migrating there and replacing Neanderthals.
In 1976 Stringer gained an ally. In that year Gunter Brauer presented the
results of his meticulous study of various archaic human remains in Africa,
such as those of Homo heidelbergensis found at Bodo, Ethiopia (fig.7)
and the more modern-looking finds at Broken Hill in Zambia. Only a few years
earlier, on the basis of associated tool-types, the Broken Hill skeletons
were still assumed to be 30,000-20,000 years old; that is, contemporary with
the Upper Paleolithic in Europe. New radiocarbon dates for the remains, however,
revealed that they were much older — some as old as 200,000 years or
more. Armed with his knowledge of these remains, Brauer developed the “out
of Africa theory,” in which he argued that modern Homo sapiens
evolved first in Africa and then migrated from there to populate the remainder
of the world. Notably, however, Brauer also allowed for some interaction
of these new humans with the native populations. For instance, he argued
that these humans were likely to have interbred with the European Neanderthals,
thus allowing for the possibility that Neanderthals had contributed somewhat
to modern populations.
Two years later, Jean-Jacques Hublin became another supporter of the replacement
model. Based on his study of the anatomy of the rear and base of Pleistocene
hominid crania, he noted that there were great morphological differences
between Neanderthal and Homo sapiens crania, and that these occured
abruptly with the introduction of the latter in Europe. He thus essentially
concurred with Stringer that modern humans could not have evolved from
Neanderthals but must have evolved outside of Europe.
In 1983, Erik
Trinkaus suggested a possible mechanism for Homo sapiens’ replacement
of Neanderthals. Analyzing several pelvic fragments from Shanidar Cave, he
suggested that Neanderthals had a larger birth canal than humans, possibly
indicating that Neanderthal babies were larger — with larger brains
— upon birth. Larger brains, however, would hav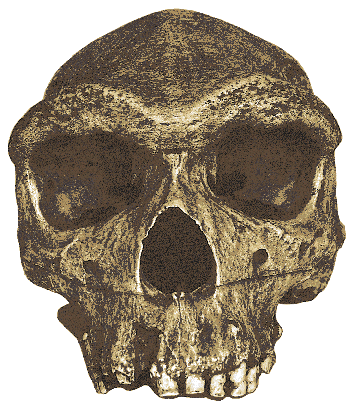 e taken longer to grow
in utero, suggesting that Neanderthals had a longer gestation period, perhaps
up to a year. If this were true, then modern humans may have had the advantage
of shorter birth spacing, thus outbreeding their Neanderthal neighbors. Although
hotly debated, the theory was discarded rather quickly when results of the
discovery of new Neanderthal remains from Kebara cave in Israel, came to
light. This first nearly complete Neanderthal pelvis, although of a male,
suggested that Neanderthal birth canals were no larger than those of modern
humans. e taken longer to grow
in utero, suggesting that Neanderthals had a longer gestation period, perhaps
up to a year. If this were true, then modern humans may have had the advantage
of shorter birth spacing, thus outbreeding their Neanderthal neighbors. Although
hotly debated, the theory was discarded rather quickly when results of the
discovery of new Neanderthal remains from Kebara cave in Israel, came to
light. This first nearly complete Neanderthal pelvis, although of a male,
suggested that Neanderthal birth canals were no larger than those of modern
humans.
Fig.7: Homo heidelbergensis skull from Bodo, Ethiopia (Athena
Review, from cast at AMNH).
As Trinkaus was revealing new insights into Neanderthal anatomy and behavior
from the Shanidar remains, new dating techniques were radically altering
the picture of other Near Eastern early human remains. The first hint that
Near Eastern hominid dating assumptions were incorrect came from Eitan
Tchernov’s 1981 study of small mammals from different levels at various
local Middle Palaeolithic sites. He found that rodents associated with the
Qafzeh remains were of archaic types similar to those from Lower Palaeolithic
levels at Tabun (i.e., below the Neanderthal remains). Therefore, Tchernov
suggested that the archaic humans from Qafzeh were older than the Neanderthals
from the other Israeli sites.
New dating techniques, such as thermoluminescence and electron spin resonance
(ESR), later applied to the same remains confirmed Tchernov's conclusions.
While Neanderthal remains from Kebara and Amud were dated to ca. 60,000 and
ca. 40,000 years ago respectively, modern humans from Qafzeh and Skhul were
dated to ca. 90,000 and ca. 80,000 years ago
(see Shea this issue).
Other dating methods indicated that the anatomically modern humans could
even be as old as 130,000 years, with the two groups overlapping in the region
for 60,000 years or more as Neanderthals moved south from Europe in accordance
with colder climactic shifts.
As these dates were creating new challenges for palaeoanthropologists, new
evidence from an unexpected field further stirred the great debate. Three
molecular biologists at the University of California at Berkeley published
a 1987 paper in Nature based on their study of mitochondrial DNA in
modern human populations, claiming a relatively recent and common origin
for all modern humans (Cann, Stoneking, and Wilson 1987). The study was based
on the presumed fact that the DNA within an intracellular structure called
mitochondria is inherited from the mother only (for other, contrary evidence
see AR 2,2 p.5), unlike the DNA from the cell's nucleus, which comes from
both parents. Therefore, mtDNA is passed down from mother to daughter, generation
to generation, without being broken up or combined, except for the natural
accumulation of mutations. By studying the mtDNA from a large group of
individuals from populations around the world they were able to show that
the number of mutations that differentiated the mtDNA of different racial
groups was very small, thus indicating that the different races must have
separated relatively recently. In addition, the biologists used a statistical
method to create an evolutionary tree showing modern humans first evolved
in Africa and then migrated from there to populate the rest of the world.
Finally, using a rate of mutation based on the date of the first migration
of modern humans to Australia and New Guinea, the researchers were actually
able to calculate the approximate date of when Homo sapiens — or
specifically a group of Homo sapiens including our ancestral
“Mitrochondrial Eve” — first evolved, sometime between 280,000
and 140,000 years ago.
Obviously this study was welcomed by proponents of the “out of Africa”
theory, but it did not go unchallenged. Critics pointed out flaws not only
in the original sample of individuals used for the study, but also in the
theoretical and statistical methods used to create the phylogenies, or
evolutionary trees. Antagonists to the genetic studies raised several key
issues, such as the effect of bottlenecking, or the possibility of an extended
period of time without change in DNA makeup. They also noted that the
evolutionary clocks, which are the basis for chronological interpretation
of DNA, are built upon assumptions of the DNA’s rate of change, potentially
rendering any subsequent timeline inaccurate.
Ten years after the original mtDNA study, new research published by Svante
Pääbo and his team appeared to confirm the replacement scenario.
The DNA for this study was retrieved directly from the original Neanderthal
from Feldhofer Cave in Germany. Comparisons of this DNA with modern human
populations suggested Neanderthals were an entirely different species from
Homo sapiens, with little or no interbreeding between the two. The study
also pointed to a date of 690,000-550,000 years ago for the split between
Neanderthals and modern human lineages. These results were essentially duplicated
in the study of DNA from a Neanderthal infant from Mezmaiskaya cave Russia.
(Ovchinnikov et al. 2000, and this issue).
More recently, however, a comprehensive analysis by Allen Templeton (2002)
of ten genetic haplotypes indicates that there have been at least three major
human migrations out of Africa (one at 1.7 mya with Homo erectus spreading
to Eurasia and Southern Asia; a second between 840,000 and 420,000 years
ago (a period that includes the evolution of archaic Homo sapiens
including Neanderthals), and the third between 150,000 and 80,000 years ago,
the time of the proposed migration by "Mitochondrial Eve's" descendants.
Yet Templeton also found there has been considerable intermixing between
the various populations used in the haplotype studies, which seem to retain
traces even of early genomes. From a genetic standpoint, this appears to
refute any complete "replacement" scenarios, including that which has been
applied to Neanderthals, and may effectively reinstate a kind of "candelabra"
model of ongoing genetic intermixing after the original migration of Homo
erectus from Africa (fig.5).
These more recent molecular studies, for all their quantitative appearance
of certainty, have not put a damper on the debates surrounding the fate of
the Neanderthals or their relationship to modern humans. While many
palaeoanthropologists hold that Homo sapiens did in some way replace
their closest cousins, how this occurred is much disputed, with some claiming
that out-and-out violence between the two groups was a factor, others arguing
that competition for resources was more important, and still others proposing
that interbreeding and “genetic swamping” was to blame for their
demise.
Additional evidence has been drawn into the debate by
Zilhăo (Duarte
et al. 1999; see also this
issue), with the discovery in November, 1998 of the complete skeleton
of a four-year-old child at the Lagar Velho rock shelter in the Lapedo Valley
of Portugal. While large parts of the site had been bulldozed, a remnant
of the original deposit contained charcoal, animal remains, and stone tools
typical of the Solutrean Period of the Upper Palaeolithic (ca. 22,000-20,000
BP). The child burial was found in the intact deposits lying 2.5 m below
this hanging remnant. Many features of the burial characterized it as a typical
human burial of the Gravettian Period of the Upper Paleolithic (ca. 27,000-22,000
BP) and it was indeed dated by radiocarbon to between 25,000-24,500 BP. Certain
characteristics of the child's anatomy, however, have led Erik Trinkaus,
who studied the remains, to propose it presented a mosaic of Neanderthal
and modern human traits. These features include Neanderthal-like limb proportions
in addition to a characteristically modern protruding chin. Based on these
findings, the excavator Joăo Zilhăo and Trinkaus have suggested
the child represents the genetic mixing of Neanderthal and modern human
populations.
Neither the Lapedo Valley child nor the new mtDNA evidence has gone unchallenged.
Debate on the relationship between modern humans and Neanderthals continues
with many basic questions unanswered. We have more sophisticated means of
evaluating Neanderthal anatomy, and no longer picture our closest human relatives
as shuffling, bent-kneed, slack-jawed brutes. Yet scholars still do not agree
on how similar Neanderthals were to anatomically modern humans in terms of
cognitive ability and behavior. Were Neanderthals capable of symbolic thought?
Did they have language, rituals and culture? Were they innovators, or merely
imitators of their more quick-brained cousins?
Closely related to these questions is how we place Neanderthals in consideration
of human origins. Could they have been merely another race in the hugely
variable and polytypic species known as Homo sapiens? Or were they an entirely
separate species, not capable of interbreeding with our own? And what was
the nature of interaction between humans and Neanderthals? We know that in
areas such as the Levant they must have lived in relatively close proximity
for thousands of years, and it is hard to accept that they did not interact
on some level. But was it through love (ie., interbreeding), a mixing of
genetic material that is evidenced in the fossil record as
“hybrids,”or was it through war, the replacement of one species
by another, by violence or competition for resources? One thing certain is
that, ultimately, Neanderthals disappeared from the earth, leaving modern
humans alone to contemplate their fate.
References:
AMNH: American Museum of Natural History, New York, NY.
Cann, R. , M. Stoneking, and A. Wilson. 1987. "Mitochondrial
DNA and Human Evolution." Nature 325: 31-36
Duarte, C., J. Mauricio, P.B. Pettit, P.Souto, E. Trinkaus, H. Van
der Plict, and J. Zilhăo. 1999. "The Early Upper Paleolithic Human
skeleton from the Abrigo do Lagar Velho (Portugal) and Modern Human Emergence
in Iberia." Proceedings of the National Academy of Sciences,
96: 7604-7609.
Huxley, T.H. 1863. On Some Fossil Remains of Man.
Krings, M. Geisert, H. Schmitz, R.W., Krainitzki, H. and Paabo, S.
1999. "DNA Sequence of the mitochondrial hypervariable region II from the
Neandertal type specimen." Proceedings of the National Academy of
Sciences, 96: 5581-5585.
Ovchinnikov, I.V, Gotherstrom, A., Romanova, G.P., Kharitonov, V.M.,
Liden, K., and Goodwin, W. 2000. "Molecular analysis
of Neanderthal DNA from the northern Caucasus." Nature 404:
490-493.
Templeton, A. 2002. "Out of Africa again and
again." Nature 416: 45-51.
Trinkaus, E., and P. Shipman. 1993. The Neandertals:
Changing the Image of Mankind. New York, Knopf.
.
This article appears in Vol.2, No.4 of Athena
Review.
.
|
|